1. Вентиляция легких. Вентиляция кровью легких. Физиологическое мертвое пространство. Альвеолярная вентиляция.
2. Перфузия легких кровью. Влияние гравитации на вентиляцию легких. Влияние гравитации на перфузию легких кровью.
3. Коэффициент вентиляционно-перфузионных отношений в легких. Газообмен в легких.
4. Состав альвеолярного воздуха. Газовый состав альвеолярного воздуха.
5. Напряжение газов в крови капилляров легких. Скорость диффузии кислорода и углекислого газа в легких. Уравнение Фика.
7. Сродство гемоглобина к кислороду. Изменение сродства гемоглобина к кислороду. Эффект Бора.
8. Углекислый газ. Транспорт углекислого газа.
9. Роль эритроцитов в транспорте углекислого газа. Эффект Холдена..
10. Регуляция дыхания. Регуляция вентиляции легких.
Кровообращение выполняет одну из важнейших функций переноса кислорода от легких к тканям, а углекислого газа - от тканей к легким. Потребление кислорода клетками тканей может изменяться в значительных пределах, например при переходе от состояния покоя к физической нагрузке и наоборот. В связи с этим кровь должна обладать большими резервами, необходимыми для увеличения ее способности переносить кислород от легких к тканям, а углекислый газ в обратном направлении.
Транспорт кислорода.
При 37 С растворимость 02 в жидкости составляет 0,225 мл л-1 кПа-1 (0,03 мл/л/мм рт. ст.). В условиях нормального парциального давления кислорода в альвеолярном воздухе, т. е. 13,3 кПа или 100 мм рт.ст., 1 л плазмы крови может переносить только 3 мл 02, что недостаточно для жизнедеятельности организма в целом. В покое в организме человека за минуту потребляется примерно 250 мл кислорода. Чтобы тканям получить такое количество кислорода в физически растворенном состоянии, сердце должно перекачивать за минуту огромное количество крови. В эволюции живых существ проблема транспорта кислорода была более эффективно решена за счет обратимой химической реакции с гемоглобином эритроцитов. Кислород переносится кровью от легких к тканям организма молекулами гемоглобина, которые содержатся в эритроцитах.
Гемоглобин способен захватывать кислород из альвеолярного воздуха (соединение называется ок-сигемоглобином) и освобождать необходимое количество кислорода в тканях. Особенностью химической реакции кислорода с гемоглобином является то, что количество связанного кислорода ограничено количеством молекул гемоглобина в эритроцитах крови. Молекула гемоглобина имеет 4 места связывания с кислородом, которые взаимодействуют таким образом, что зависимость между парциальным давлением кислорода и количеством переносимого кислорода с кровью имеет S-образную форму, которая носит название кривой насыщения или диссоциации оксигемоглобина (рис. 10.18). При парциальном давлении кислорода 10 мм рт. ст. насыщение гемоглобина кислородом составляет примерно 10 %, а при Р02 30 мм рт. ст. - 50-60 %. При дальнейшем увеличении парциального давления кислорода от 40 мм рт. ст. до 60 мм рт. ст. происходит уменьшение крутизны кривой диссоциации оксигемоглобина и процент его насыщения кислородом возрастает в диапазоне от 70-75 до 90 % соответственно. Затем кривая диссоциации оксигемоглобина начинает занимать практически горизонтальное положение, поскольку увеличение парциального давления кислорода с 60 до 80 мм рт. ст. вызывает прирост насыщения гемоглобина кислородом на 6 %. В диапазоне от 80 до 100 мм рт. ст. процент образования оксигемоглобина составляет порядка 2. В результате кривая диссоциации оксигемоглобина переходит в горизонтальную линию и процент насыщения гемоглобина кислородом достигает предела, т. е. 100. Насыщение гемоглобина кислородом под влиянием Р02 характеризует своеобразный молекулярный «аппетит» этого соединения к кислороду.
Значительная крутизна кривой насыщения гемоглобина кислородом в диапазоне парциального давления от 20 до 40 мм рт. ст. способствует тому, что в ткани организма значительное количество кислорода может диффундировать из крови в условиях фадиента его парциального давления между кровью и клетками тканей (не менее 20 мм рт. ст.). Незначительный процент насыщения гемоглобина кислородом в диапазоне его парциального давления от 80 до 100 мм рт. ст. способствует тому, что человек без риска снижения насыщения артериальной крови кислородом может перемещаться в диапазоне высот над уровнем моря до 2000 м.
 Рис. 10.18. Кривая диссоциации оксигемоглобина
. Пределы колебания кривой при РС02 = 40 мм рт. ст. (артериальная кровь) и РС02 = 46 мм рт. ст. (венозная кровь) показывают изменение сродства гемоглобина к кислороду (эффект Ходена
).
Рис. 10.18. Кривая диссоциации оксигемоглобина
. Пределы колебания кривой при РС02 = 40 мм рт. ст. (артериальная кровь) и РС02 = 46 мм рт. ст. (венозная кровь) показывают изменение сродства гемоглобина к кислороду (эффект Ходена
).
Общие запасы кислорода в организме обусловлены его количеством, находящимся в связанном состоянии с ионами Fe2+ в составе органических молекул гемоглобина эритроцитов и миоглобина мышечных клеток.
Один грамм гемоглобина связывает 1,34 мл 02. Поэтому в норме при концентрации гемоглобина 150 г/л каждые 100 мл крови могут переносить 20,0 мл 02.
Количество 02, которое может связаться с гемоглобином эритроцитов крови при насыщении 100 % его количества, называется кислородной емкостью гемоглобина . Другим показателем дыхательной функции крови является содержание 02 в крови (кислородная емкость крови ), которое отражает его истинное количество, как связанного с гемоглобином, так и физически растворенного в плазме. Поскольку в норме артериальная кровь насыщена кислородом на 97 %, то в 100 мл артериальной крови содержится примерно 19,4 мл 02.
Для начала восстановим в памяти несколько положений и понятий из области физики, без них изучение газообмена и транспорта газа в легких, невозможно. Итак, атмосферный воздух имеет довольно постоянный процентный состав газов. Это завидное постоянство характерно и для альвеолярного воздуха, то есть для того, который не просто заполняет легкие, а контактирует непосредственно с пневмоцитами, выстилающими альвеолы. Правда, О2 в альвеолярном воздухе меньше, чем его содержание в атмосферном (14 и 21%, соответственно), а СО2 значительно больше (5,5 против 0,03% в атмосферном), но значения эти (14 и 5,5%) постоянные (разница между альвеолярным и атмосферным воздухом - результат постоянно происходящего газообмена, находящегося вне зависимости от времени суток, а также от того вдох сейчас или выход, хочет того человек или нет).
А теперь вводим первое физическое понятие - парциальное давление газа . В воздухе, представленном в виде газовой смеси оно пропорционально процентному содержанию этого газа в общем давлении смеси. Атмосферное давление, как известно, равняется 760 мм рт.ст. Давление же газовой смеси в альвеолярном воздухе несколько меньше, так как часть его пришлась на возрастающее в дыхательной системе количество водяных паров, и составляет 713 мм рт.ст. Теперь не составит труда простыми пропорциями рассчитать парциальное давление в альвеолярном воздухе кислорода и углекислого газа. Если давление газовой смеси 713 мм рт.ст., а кислорода содержится 14%, значит парциальное давление О2 равно 100 мм рт.ст. Так же находим это значение и для углекислоты - оно будет равно 40 мм рт.ст. Стоит запомнить, что парциальное давление обоих газов в альвеолярном воздухе являет собой ту силу, с которой молекулы этих газов пытаются проникнуть через аэрогематинеский барьер в кровь из альвеол легких.
Что же мешает такому переходу? Оказывается, в плазме крови этих газов и без того достаточно. Они находятся там в растворенном виде, и, мало того, сами не прочь выйти из раствора в альвеолярный воздух. Здесь влияние оказывает напряжение газа , находящегося в жидкости. Напряжение газа - это величина, характеризующая силу стремления молекул растворенного газа выйти из водной среды в газовую. В физическом отношении понятия «парциальное давление» и «напряжение» очень близки, только относятся к разным средам: первое - к газовой смеси, а второе - к жидкости. Но самое главное в том, что они противостоят друг другу. Если бы парциальное давление, скажем, СО2 было равно напряжению СО2 в крови, то передвижение двуокиси углерода не наблюдалось бы ни в том, ни в другом направлении.
И все же газообмен происходит. И происходит он благодаря разнице характеристик парциального давления газов, находящихся в альвеолярном воздухе с напряжением тех же самых газов, находящихся в плазме крови. Взгляните на рисунок справа. Начнем с кислорода. К легким по системе легочной артерии притекает кровь, бедная О2, напряжение которого в ней равно 40 мм рт.ст. Кровь бежит по капиллярам, расположенным в межальвеолярных стенках, то есть через аэрогематический барьер соседствует с воздухом альвеол, в котором парциальное давление О2 равно 100 мм рт.ст. То есть мы наблюдаем разницу 40 и 100! Конечно, О2 устремляется в кровь и он будет растворятся в крови до тех самых пор, пока напряжение этого газа не увеличится хотя бы до 96 мм рт.ст. Когда артериальная кровь насыщается кислородом, то собирается в легочных венах, чтобы через них покинуть легкие.
Другая ситуация складывается с CO2. Кровь, поступает к легким от всего организма через сосуды малого круга, она содержит много СО2 (46 мм рт.ст.), однако парциальное давление СО2 в альвеолах только 40 мм. Это и определяет движение двуокиси углерода из плазмы через барьер для последующего высвобождения в альвеолярный воздух, что приводит к снижению напряжения СО2 до 39 мм рт.ст.
За транспорт кислорода от легких к тканям в основном отвечают эритроциты. Когда в легочных капиллярах начинает нарастать напряжение кислорода, гемоглобин эритроцитов начинает выхватывать из плазмы молекулы О2, постепенно превращаясь в оксигемоглобин. Именно в такой форме кислород приносится к органам и тканям. Оксигемоглобин «отсоединяет» от себя О2, отдавая его снова в плазму, и начинается как бы вторая серия - газообмен осуществляется уже между кровью и тканями.
Всем клеткам организма нужен кислород, т.к. именно этот газ является универсальным окислителем в процессах. Используя кислород в биохимических реакциях, клетки получают необходимую им энергию и углекислый газ, требующий удаления за пределы клетки. Так как не все клетки оказываются в непосредственном контакте с капиллярами, то надежным посредником между ними является тканевая жидкость, о которой подробнее будет рассказано в параграфах о внутренней среде организма и о лимфе. Из тканевой жидкости клетка забирает кислород, поступающий из капилляра, в нее же «выбрасывает» двуокись углерода. Другими словами, тканевой газообмен осуществляется главным образом между плазмой крови и жидкостью тканей организма. А там уже все проходит по известному механизму. Еще раз обратитесь к таблице на рис. 66. Напряжение О2 в тканевой жидкости невелико (40 мм рт.ст.), чего не скажешь о крови артерий (96 мм рт.ст.). Исходя из этого столь необходимый клеткам кислород перемещается из плазмы в тканевую жидкость до тех пор, пока напряжение этого газа в крови не достигнет 40 мм рт.ст. СО2 же газ из места большего своего напряжения (46 мм рт.ст. в жидкости тканей) устремляется в плазму крови, где его напряжение составляет 39 мм рт.ст., доводя его до отметки 46. Кровь с такими показателями О2 и СО2 (40 мм и 46 мм рт.ст.) будет венозной и по венам большого круга притекает к правым отделам сердца, откуда отправляется для осуществления газообмена в легкие человека.
Транспорт углекислого газа в организме человека способен осушествлятся кровью 3-мя путями. Незначительная часть газа растворяется в плазме, определяя тем самым напряжение СО2 в крови. Большая часть СО2 тем временем контактирует с гемоглобином красных кровяных телец, соединяется с ним, превращая в карбоксигемоглобин. Ну а весь оставшийся СО2 транспортируется в качестве кислых солей угольной кислоты (чаще всего NaHCO3). Тем ни менее, какой бы способ транспортировки углекислоты не использовался, он приводит газ к легких, для последующего его выведения из человеческого организма.
Итак, если постараться кратко резюмировать, то можно сказать, что существует 2 стадии газообмена : легочная и тканевая. В легочной стадии основой считается разница парциального давления газа в альвеолярном воздухе с напряжением газа в крови. Для тканевой стадии основой будет разница напряжения газа в крови и жидкости тканей. Сам транспорт газов обязательно происходит, если газы находятся в растворенном виде, либо в связанном, если молекулы газов соединяются с ионами или молекулой гемоглобина.
Транспорт кислорода кровью. Кислород транспортируется кровью в двух формах - в растворенном виде и в соединении с гемоглобином. В плазме артериальной крови содержится очень небольшое количество физически растворенного кислорода, всего
0,3 об.%, т. е. 0,3 мл кислорода в 100 мл крови. Основная же часть кислорода вступает в непрочное соединение с гемоглобином эритроцитов, образуя оксигемоглобин. Насыщение крови кислородом называется оксигенацией или артериализацией крови. Кровь, оттекающая от легких по легочным венам, имеет такой же газовый состав, что и артериальная кровь в большом круге кровообращения.
Количество кислорода, находящееся в 100 мл крови при условии полного перехода гемоглобина в оксигемоглобин, называется кислородной емкостью крови. Эта величина помимо парциального давления кислорода зависит от содержания гемоглобина в крови. Известно, что 1 г гемоглобина может в среднем связать 1,34 мл кислорода. Следовательно, зная уровень содержания в крови гемоглобина, можно вычислить кислородную емкость крови. Так, у лошадей при содержании гемоглобина в крови около 14 г/100 мл кислородная емкость крови составляет (1,34 14) около 19 об.%, у крупного рогатого скота при уровне гемоглобина 10... 12 г/100 мл - около 13...16 об.%. Пересчитав содержание кислорода в общем объеме крови, оказывается, что его запаса хватит лишь на З...4мин при условии, если он не будет поступать из воздуха.
На уровне моря при соответственных колебаниях атмосферного давления и парциального давления кислорода в альвеолярном воздухе гемоглобин практически полностью насыщается кислородом. В условиях высокогорья, где атмосферное давление низкое, снижается парциальное давление кислорода и уменьшается кислородная емкость крови. На содержание кислорода в крови также влияет температура крови: с повышением температуры тела снижается кислородная насыщенность крови. Высокое содержание в крови водородных ионов и диоксида углерода способствует отщеплению кислорода от оксигемогло-бина при прохождении крови через капилляры большого круга кровообращения.
Обмен газов между кровью и тканями совершается так же, как и обмен газов между кровью и альвеолярным воздухом - по законам диффузии и осмоса. Поступающая сюда артериальная кровь насыщена кислородом, его напряжение составляет 100 мм рт. ст. В тканевой жидкости напряжение кислорода составляет 20...37 мм рт. ст., а в клетках, которые потребляют кислород, его уровень падает до 0. Поэтому оксигемоглобин отщепляет кислород, который переходит сначала в тканевую жидкость, а затем в клетки тканей.
В процессе тканевого дыхания из клеток выделяется диоксид углерода. Он сначала растворяется в тканевой жидкости и создает там напряжение около 60...70 мм рт. ст., что выше, чем в крови (40 мм рт. ст.). Градиент напряжения кислорода в тканевой жидкости и крови является причиной диффузии диоксида углерода из тканевой жидкости в кровь.
Транспорт диоксида углерода кровью. Диоксид углерода транспортируется в трех формах: в растворенном виде, в соединении с гемоглобином (карбогемоглобин) и в виде бикарбонатов.
Поступающий из тканей диоксид углерода незначительно растворяется в плазме крови -до 2,5об.%; его растворимость немного выше, чем у кислорода. Из плазмы диоксид углерода проникает в эритроциты и вытесняет из оксигемоглобина кислород. Оксигемоглобин превращается в восстановленнный, или редуцированный, гемоглобин. Присутствующий в эритроцитах фермент 4 карбоангидраза ускоряет соединение диоксида углерода с водой и образование угольной кислоты - Н 2 С0 3 . Эта кислота нестойкая, она диссоциирует на Н + и HCOJ.
Поскольку мембрана эритроцита непроницаема для Н + , он остается в эритроцитах, а НС0 3 переходит в плазму крови, где превращается в бикарбонат натрия (NaHC0 3). Часть диоксида углерода в эритроцитах соединяется с гемоглобином, образуя карбогемоглобин, а с катионами калия - бикарбонат калия (КНС0 3).
В легочных альвеолах, где парциальное давление диоксида углерода ниже, чем в венозной крови, растворенный и освободившийся при диссоциации карбогемоглобина диоксид углерода диффундирует в альвеолярный воздух. Одновременно кислород переходит в кровь и связывается с редуцированным гемоглобином, образуя оксигемоглобин. Оксигемоглобин, являясь более сильной кислотой, чем угольная, вытесняет угольную кислоту из бикарбонатов ионы калия. Угольная кислота расщепляется до С0 2 и Н 2 0 при участии карбоангидразы. Диоксид углерода переходит из эритроцитов в плазму крови и затем в альвеолярный воздух (см. рис. 7.6).
Несмотря на то что основная часть диоксида углерода присутствует в плазме крови в форме бикарбоната натрия, в альвеолярный воздух выделяется преимущественно диоксид углерода не из плазмы крови, а из эритроцитов. Дело в том, что только в эритроцитах имеется карбоангидраза, расщепляющая угольную кислоту. В плазме крови карбоангидразы нет, поэтому бикарбонаты разрушаются очень медленно и диоксид углерода не успевает выйти в альвеолярный воздух (по легочным капиллярам кровь проходит менее чем за 1 с). Таким образом, диоксид углерода находится в крови в трех формах: растворенной, в виде карбогемоглобина, бикарбонатов, но через легкие удаляется только в одной форме - С0 2 .
Не весь кислород из артериальной крови поступает в ткани, часть его переходит в венозную кровь. Отношение объема кислорода, поглощенного тканями, к содержанию его в артериальной крови называется коэффициентом утилизации кислорода. В условиях физиологического покоя он составляет около 40 %. При более высоком уровне метаболизма коэффициент утилизации кислорода увеличивается и уровень его в венозной крови падает.
Проходя через легкие, не весь диоксид углерода поступает в альвеолярный воздух, часть его остается в крови и переходит в арте-
риальную кровь. Таким образом, если в венозной крови содержится 58 об.% диоксида углерода, то в артериальной крови - 52 об.%. Наличие определенного уровня кислорода и особенно диоксида углерода в артериальной крови имеет огромное значение в процессах регуляции внешнего дыхания.
Тканевое (внутриклеточное) дыхание. Тканевое дыхание - это процесс биологического окисления в клетках и тканях организма.
Биологическое окисление происходит в митохондриях. Внутреннее пространство митохондрий окружено двумя мембранами-наружной и внутренней. На внутренней мембране, имеющей складчатое строение, сосредоточено большое количество ферментов. Поступающий в клетку кислород затрачивается на окисление жиров, углеводов и белков. При этом освобождается энергия в наиболее доступной для клеток форме, прежде всего в форме АТФ - аденозинтрифосфорной кислоты. Ведущее значение в окислительных процессах имеют реакции дегидрирования (отдача водорода).
Синтез АТФ осуществляется при миграции электронов от субстрата к кислороду через цепь дыхательных ферментов (фла-виновые ферменты, цитохромы и др.) Освобождающаяся энергия накапливается в форме макроэргических соединений (например, АТФ), а конечными продуктами реакций становятся вода и диоксид углерода.
Наряду с окислительным фосфорилированием кислород может использоваться в некоторых тканях по типу непосредственного внедрения в окисляемое вещество. Такое окисление называется микросомальным, ибо происходит в микросомах - везикулах, образованных мембранами эндоплазматического ретику-лума клетки.
Ткани и органы имеют разную потребность в кислороде: интенсивнее поглощают кислород из крови головной мозг, особенно кора больших полушарий, печень, сердце, почки. Меньше потребляют кислорода в состоянии покоя клетки крови, скелетные мышцы, селезенка. При нагрузке потребление кислорода возрастает. Например, при тяжелой мышечной работе скелетные мышцы потребляют больше кислорода в 40 раз, сердечная мышца - в 4 раза (в расчете на 1г ткани).
Даже в пределах одного органа потребление кислорода может резко отличаться. Например, в корковой части почек оно интенсивнее, чем в мозговой части, в 20 раз. Это зависит от строения ткани, плотности распределения в ней кровеносных капилляров, регуляции кровотока, коэффициента утилизации кислорода и ряда других факторов. Следует помнить, что чем больше клетки будут потреблять кислорода, тем больше образуется продуктов обмена - диоксида углерода и воды.
7.4. РЕГУЛЯЦИЯ ДЫХАНИЯ
Главная биологическая функция дыхания - это обеспечение газообмена в тканях. Именно ради тканевого дыхания в процессе эволюции возникли и совершенствовались системы кровообращения и внешнего дыхания. Доставка тканям кислорода, выведение водородных ионов и диоксида углерода должны точно соответствовать потребностям тканей и организма в определенный период их жизнедеятельности. В реализации этих процессов, их динамического равновесия участвуют сложные механизмы, включающие регуляцию газового состава крови, регионального кровообращения и трофику тканей. В данной главе мы рассмотрим, каким образом организм поддерживает определенное содержание кислорода и диоксида углерода в крови, т. е. каким образом регулируются дыхательные циклы, глубина и частота дыхания.
Внешнее дыхание регулируется нейрогуморальными механизмами. Еще в 1885 г. русский физиолог Н. А. Миславский обнаружил в продолговатом мозге дыхательный центр и доказал наличие в нем двух отделов - центр вдоха и центр выдоха. Центробежными (эфферентными) путями дыхательный центр связан с эффекторами - дыхательными мышцами. Афферентные, или сенсорные, или центростремительные, импульсы поступают в дыхательный центр от различных экстеро- и интерорецепторов, а также от вышележащих отделов головного мозга. Таким образом, в виде довольно упрощенной общей схемы можно представить типичную рефлекторную дугу, состоящую из рецепторов, афферентных путей, нервного центра, эфферентных путей и эффекторов - дыхательных мышц.
Дыхательный центр. Дыхательный центр - это совокупность нейронов, расположенных во всех отделах ЦНС и принимающих то или иное участие в регуляции дыхания. Главная часть, или как принято говорить, «ядро» дыхательного центра, находится, как доказал экспериментально Миславский, в продолговатом мозге, в области ретикулярной формации на дне четвертого мозгового желудочка. Без этого отдела дыхание невозможно, повреждение продолговатого мозга приводит неминуемо к смерти из-за остановки дыхания.
Четкий морфологический раздел между центрами вдоха и выдоха в продолговатом мозге отсутствует, но существует распределение функций между нейронами: одни нейроны - инспиратор-ные - генерируют потенциалы действия, возбуждающие инспи-раторные мышцы, другие - экспираторные - возбуждают выдыхательные мышцы.
В инспираторных нейронах электрическая активность включается быстро, постепенно нарастает частота им-пульсации (до 70... 100 импульсов в 1 с) и резко падает к концу вдоха. Эта импульсация приводит к сокращению диафрагмы,
межреберных и других инспираторных мышц. «Выключение» инспираторных нейронов приводит к расслаблению инспираторных мышц и выдоху. Активность экспираторных нейронов при нормальном спокойном дыхании имеет меньшее значение. Но при усиленном дыхании, особенно при форсированном выдохе, экспираторные нейроны определяют сокращение выдыхательных мышц.
Буль барный отдел дыхательного центра головного мозга обладает автоматией. Эта уникальная особенность дыхательного центра заключается в том, что его нейроны могут спонтанно, т. е. самопроизвольно, без каких-либо внешних воздействий деполяризовываться, или разряжаться. Впервые спонтанные колебания электрической активности дыхательного центра обнаружил И. М. Сеченов. Природа автоматии дыхательного центра до сих пор не выяснена. Вероятно, она зависит от специфики обмена веществ нейронов этой области головного мозга и особой чувствительности инспираторных нейронов к окружающей среде, составу цереброспинальной жидкости. Автоматия дыхательного центра сохраняется после почти полной его деаффе-рентации, т. е. после прекращения воздействий со стороны различных рецепторов.
Благодаря автоматии дыхательный центр продолговатого мозга обеспечивает ритмичные чередования вдоха и выдоха и определяет частоту дыхания в условиях физиологического покоя.
Бульбарный отдел дыхательного центра является самым устойчивым отделом ЦНС к действию наркотических препаратов. Даже при глубоком наркозе, когда рефлекторные реакции отсутствуют, самостоятельное дыхание сохраняется. В арсенале фармакологических препаратов имеются вещества, избирательно повышающие возбудимость дыхательного центра, - лобелии, цититон, воздействующие на дыхательный центр рефлекторно, через рецепторы синокаротидной зоны.
И. П. Павлов говорил, что дыхательный центр, который раньше представляли величиной с булавочную головку, необычайно разросся: он спустился вниз в спинной мозг и поднялся вверх до коры больших полушарий.
Какую же роль играют другие отделы дыхательного центра? В спинном мозге находятся нейроны (мотонейроны), иннерви-руюшие дыхательные мышцы (рис. 7.7). Возбуждение к ним передается от инспираторных и экспираторных нейронов продолговатого мозга по нисходящим проводящим путям, лежащим в белом веществе спинного мозга. В отличие от бульварного центра мотонейроны спинного мозга не обладают автоматией. Поэтому после перерезки спинного мозга сразу за продолговатым дыхание останавливается, так как дыхательные мышцы не получают команды к сокращениям. Если же спинной мозг перерезать на уровне 4...5-го шейного позвонка, то самостоятельное
Рис. 7.7. Схема организации центрального аппарата регуляции дыхания
дыхание сохраняется за счет сокращений диафрагмы, потому что центр диафрагмального нерва расположен в 3...5-M шейных сегментах спинного мозга.
Выше продолговатого мозга, прилегая к нему, находится варолиев мост, в котором расположен «пневмотаксический центр». Он не обладает автоматией, но благодаря непрерывной активности обеспечивает периодическую деятельность дыхатель-
ного центра, увеличивает скорость развития инспираторной и экспираторной импульсаций в нейронах продолговатого мозга.
Средний мозг имеет большое значение в регуляции тонуса поперечнополосатых мышц. Поэтому при сокращении различных мышц афферентная импульсация от них поступает в средний мозг, который соответственно мышечной нагрузке изменяет характер дыхания. Средний мозг ответствен также за координацию дыхания с актами глотания, рвоты и отрыгивания. Во время глотания дыхание задерживается на фазе выдоха, надгортанник закрывает вход в гортань. При рвоте, отрыгивании газов происходит «холостой вдох» - вдох при закрытой гортани. При этом сильно снижается внутриплевральное давление, что и способствует поступлению содержимого из желудка в грудную часть пищевода.
Гипоталамус - отдел промежуточного мозга. Значение гипоталамуса в регуляции дыхания заключается в том, что в нем содержатся центры, контролирующие все виды обмена веществ (белковый, жировой, углеводный, минеральный), и центр тепло-регуляции. Поэтому усиление обмена веществ, повышение температуры тела ведут к усилению дыхания. Например, при повышении температуры тела дыхание учащается, что способствует увеличению отдачи теплоты вместе с выдыхаемым воздухом и предохраняет организм от перегревания (тепловая одышка).
Гипоталамус принимает участие в изменении характера ды
хания при болевых раздражениях, при различных поведенчес
ких актах (прием корма, обнюхивание, спаривание и др.). По
мимо регуляции частоты и глубины дыхания гипоталамус через
вегетативную нервную систему регулирует просвет бронхиол,
спадение нефункционирующих альвеол, степень расширения
легочных сосудов, проницаемость легочного эпителия и стенок
капилляров. /
Многогранно значение коры больших полушарий головного мозга в регуляции дыхания. В коре расположены центральные отделы всех анализаторов, информирующих как о внешних воздействиях, так и о состоянии внутренней среды организма. Поэтому наиболее тонкое приспособление дыхания к сиюминутным потребностям организма осуществляется при обязательном участии высших отделов нервной системы.
Особое значение имеет кора больших полушарий при мышечной работе. Известно, что учащение дыхания начинается за несколько секунд до начала работы, сразу после команды «приготовиться». Аналогичное явление наблюдается у спортивных лошадей наряду с тахикардией. Причиной подобных «опережающих» реакций у людей и животных являются выработавшиеся в результате повторных тренировок условные рефлексы. Только влиянием коры больших полушарий можно объяснить произвольные, волевые изменения ритма, частоты и глубины дыхания. Человек может произвольно задержать дыхание на несколько секунд или усилить
его. Несомненна роль коры в изменении паттерна дыхания во время подачи голоса, при нырянии, обнюхивании.
Итак, в регуляции внешнего дыхания участвует дыхательный центр. Ядро этого центра, находящееся в продолговатом мозге, посылает ритмичные импульсы через спинной мозг к дыхательным мышцам. Сам же бульварный отдел дыхательного центра находится под постоянным воздействием со стороны вышележащих отделов ЦНС и различных рецепторов - пульмональных, сосудистых, мышечных и др.
Значение рецепторов легких в регуляции дыхания. В легких имеются три группы рецепторов: растяжения и спадения; ирритантные; юкстакапиллярные.
Рецепторы растяжения расположены между гладкими мышцами в воздухоносных путях - вокруг трахеи, бронхов и бронхиол, а в альвеолах и плевре отсутствуют. Растяжение легких при вдохе вызывает возбуждение механорецепторов. Возникающие потенциалы действия передаются по центростремительным волокнам блуждающего нерва в продолговатый мозг. К концу вдоха частота импульсации нарастает от 30 до 100 импульсов в 1 с и становится пессимальной, вызывая торможение центра вдоха. Начинается выдох. Рецепторы спадения легких изучены недостаточно. Возможно, при спокойном дыхании их значение невелико.
Рефлексы с механорецепторов легких названы по имени открывших их ученых - рефлексы Геринга - Брейера. Назначение этих рефлексов заключается в следующем: информировать дыхательный центр о состоянии легких, их наполненности воздухом и в соответствии с этим регулировать последовательность вдоха и выдоха, ограничивать чрезмерное растяжение легких при вдохе или спадение легких при выдохе. У новорожденных рефлексы с механорецепторов легких играют большую роль; с возрастом значение их уменьшается.
Таким образом, значение блуждающего нерва в регуляции дыхания заключается в передаче афферентных импульсов от механорецепторов легких в дыхательный центр. У животных после перерезки вагуса информация от легких не попадает в продолговатый мозг, поэтому дыхание становится медленным, с коротким вдохом и очень продолжительным выдохом (рис. 7.8). При раздражении вагуса наблюдается задержка дыхания в зависимости от того, в какую фазу дыхательного
цикла действует раздражение. Если раздражение поступает во время вдоха, то вдох преждевременно прекращается и сменяется выдохом, а если совпадает с фазой выдоха, то, наоборот, выдох сменяется вдохом (рис. 7.9).
Ирритантные рецепторы расположены в эпителиальном и субэпителиальном слоях всех воздухоносных путей. Они раздражаются при попадании в воздухоносные пути пыли, ядовитых газов, а также при достаточно больших изменениях объема легких. Некоторая часть ирритантных рецепторов возбуждается при обычных вдохах и выдохах. Рефлексы с ирритантных рецепторов носят защитный характер - чихание, кашель, глубокий вдох («вздох»). Центры данных рефлексов расположены в продолговатом мозге.
Юкстакапиллярные рецепторы (юкста - вокруг) находятся вблизи капилляров малого круга кровообращения. По функциям они сходны с рецепторами спадения, для них раздражителем является увеличение интерстициального пространства легких, например при отеке. Раздражение юкстакапилляр-ных рецепторов вызывает одышку. Возможно, при интенсивной мышечной работе кровяное давление в легочных сосудах повышается, это увеличивает объем интерстициальной жидкости и стимулирует активность юкстакапиллярных рецепторов. Раздражителем пульмональных рецепторов может быть гистамин, синтезирующийся в базофилах и тучных клетках. В легких этих клеток довольно много, и при аллергических заболеваниях они выделяют гистамин в таком количестве, что это приводит к отеку и одышке.
Значение рецепторов дыхательных мышц. В дыхательных мышцах имеются рецепторы растяжения - мышечные веретена, сухожильные рецепторы. Особенно велика плотность их размещения в межреберных мышцах и мышцах стенок живота. Механорецепто-ры дыхательных мышц возбуждаются при их сокращении или растяжении при вдохе или выдохе. По принципу обратной связи они регулируют возбуждение мотонейронов спинного мозга в зависимости от их исходной длины и сопротивления, которое они встре-
Чают при сокращении. Сильное раздражение механорецепторов грудной клетки (например, при ее сжатии) вызывает торможение инспираторной деятельности дыхательного центра.
Значение хеморецепторов в регуляции дыхания. Исключительно важное значение в регуляции внешнего дыхания имеет газовый состав артериальной крови. Биологическая целесообразность этого вполне понятна, поскольку от содержания кислорода и диоксида углерода в артериальной крови зависит обмен газов между кровью и тканями. Давно стали классикой опыты Фредерика (1890) с перекрестным кровообращением, когда артериальная кровь от одной собаки поступала в кровь другой, а венозная кровь от головы второй собаки - в венозную кровь первой собаки (рис. 7.10). Если пережать трахею и тем самым остановить дыхание первой собаки, то ее кровь с недостаточным содержанием кислорода и избыточным диоксида углерода омывает головной мозг второй собаки. Дыхательный центр второй собаки усиливает дыхание (гипер-пноэ), и в ее крови снижается концентрация диоксида углерода и урежается дыхание вплоть до остановки (апноэ).
Благодаря опытам Фредерика стало очевидным, что дыхательный центр чувствителен к уровню содержания газов в артериальной крови. Возросшая концентрация диоксида углерода (гипер-капния) и водородных ионов в крови вызывает учащение дыхания, вследствие чего диоксид углерода выделяется с выдыхаемым воздухом и его концентрация в крови восстанавливается. Снижение содержания диоксида углерода в крови (гипокапния), напротив, вызывает урежение дыхания или его остановку до тех пор, пока в крови концентрация диоксида углерода снова не достигнет нормальной величины (нормокапния).
Концентрация кислорода в крови также влияет на возбудимость дыхательного центра, но в меньшей мере, чем диоксида углерода. Это связано с тем, что при обычных колебаниях атмосферного давления, даже на высотах до 2000 м над уровнем моря, почти весь гемоглобин превращается в оксигемоглобин, поэтому парциальное давление кислорода в артериальной крови всегда выше, чем в тканевой жидкости, и ткани получают, во всяком случае в
состоянии физиологического покоя, достаточно кислорода. При значительном снижении парциального давления кислорода в воздухе уменьшается содержание кислорода в крови (гипоксемия) и в тканях (гипоксия), в результате этого возбудимость дыхательного центра повышается и дыхание учащается.
Снижение концентрации кис-Рис. 7.10. Перекрестное кровообращение лорода В крови (гипоксемия) МО-
жет произойти и вследствие более интенсивного потребления его тканями. В этом случае возможно развитие кислородной недостаточности, что, в свою очередь, вызовет усиление внешнего дыхания. При повышении содержания кислорода в крови, например при вдыхании газовой смеси с высоким содержанием кислорода или при нахождении в барокамере под высоким атмосферном давлении, вентиляция легких уменьшается за счет угнетения дыхательного центра.
Мы рассмотрели в отдельности значение содержания кислорода и диоксида углерода в артериальной крови, т. е. аналитически. Однако в действительности оба газа влияют на дыхательный центр одновременно. Установлено, что гипоксия повышает чувствительность дыхательного центра к повышенному содержанию диоксида углерода, и усиление дыхания в этих условиях является интегральной реакцией дыхательного центра в ответ на изменение газового состава крови. Так, при физической работе в мышцы поступает больше кислорода из притекающей крови, увеличивается коэффициент утилизации кислорода, а его концентрация в крови снижается. Одновременно в результате повышения метаболизма из мышц в кровь поступает больше углекислоты и органических кислот.
Велика роль сосудистых хеморецепторов при первом вдохе новорожденного. Снижение содержания кислорода в крови и увеличение диоксида углерода во время родов, особенно после пережатия пуповины, является главнейшим раздражителем дыхательного центра, что и вызывает первый вдох.
Если в течение 1 мин произвольно максимально усилить дыхание и вызвать этим гипервентиляцию легких, то заметно удлиняется дыхательная пауза между выдохом и последующим вдохом. Может наступить кратковременное апноэ - остановка дыхания на 1...2 мин. Без предшествующей гипервентиляции задержать дыхание можно лишь на 20...30 с. Подобную гипервентиляцию легких с последующим апноэ вызывают у себя ныряльщики - охотники за жемчугом или губкой. После длительных тренировок они остаются под водой до 4...5 мин.
Попробуем разобраться в механизмах апноэ после одышки. Поскольку при обычном спокойном дыхании кровь насыщена кислородом на 95 %, усиление дыхания не приводит к значительному увеличению концентрации кислорода в крови. На содержание же диоксида углерода гипервентиляция оказывает заметное влияние - уровень диоксида углерода снижается сначала в альвеолярном воздухе, а затем в крови. Следовательно, апноэ после гипервентиляции легких связано с уменьшением концентрации углекислоты в крови. Дыхание восстановится, когда в крови снова накопится достаточный, или пороговый, уровень диоксида углерода.
Если задержать дыхание на 20...30 с, то наступает неудержимое стремление вздохнуть и сделать несколько глубоких дыхательных движений. Следовательно, задержка ведет к гиперпноэ - усиле-
нию дыхания. Это также обусловлено накоплением в крови диоксида углерода, так как за 20...30 с концентрация кислорода в крови снизится незначительно, а диоксид углерода постоянно поступает в кровь из тканей.
Итак, диоксид углерода является главнейшим гуморальным раздражителем дыхательного центра. Изменение его концентрации в крови ведет к таким изменениям в частоте и глубине дыхания, которые восстанавливают постоянный уровень углекислоты в крови. При увеличении уровня диоксида углерода в крови происходит стимуляция дыхательного центра и усиление дыхания, при снижении - уменьшение частоты и глубины дыхания. Поэтому столь эффективен метод искусственного дыхания «изо рта в рот», а в газовые смеси для искусственного дыхания обязательно добавляют диоксид углерода.
Где же находятся те датчики, или рецепторы, которые улавливают концентрацию газов в крови? Они расположены там, где необходим тщательный контроль за газовым составом внутренней среды организма. Такими участками являются сосудистые рефлексогенные зоны каротидного синуса и аорты, а также центральные рефлексогенные зоны в продолговатом мозге.
Синокаротидная зона, или зона каротидного синуса, имеет особо важное значение в отслеживании газового состава и рН крови. Она находится в области разветвления сонных артерий на наружные и внутренние ветви, откуда артериальная кровь направляется в головной мозг. Пороговая концентрация кислорода, углекислоты и водородных ионов для рецепторов синокаротидной зоны соответствует их уровню в крови при нормальных условиях в состоянии покоя. Небольшое возбуждение возникает в отдельных рецепторах при редком глубоком дыхании, когда концентрация газов в крови начинает немного изменяться. Чем сильнее изменяется газовый состав крови, тем большая частота импульсации возникает в хеморецепторах, стимулируя дыхательный центр.
Изменение дыхательных движений происходит не только при раздражении хеморецепторов аорты или каротидного синуса. Раздражение находящихся здесь же баро- или прессорецепторов при повышении артериального давления обычно ведет к замедлению дыхания, а при снижении артериального давления - к его усилению. Однако при физической нагрузке повышение артериального давления не приводит к угнетению дыхания, а также к депрессор-ным рефлексам.
Центральные (медуллярные) хеморецепторы в продолговатом мозге чувствительны к уровню содержания диоксида углерода в цереброспинальной жидкости. Если артериальные хеморецепторы регулируют газовый состав артериальной крови, то центральные хеморецепторы держат под контролем газовый и кислотно-щелочной гомеостаз жидкости, омывающей головной мозг, - наиболее уязвимую ткань организма. Хеморецепторы, чувствительные к из-
менению рН, диоксида углерода и кислорода, имеются также в венозных сосудах и в различных тканях организма. Однако их значение заключается не в регуляции внешнего дыхания, а в изменении регионального, или местного, кровотока.
Большой интерес представляют механизмы изменения дыхания при физической работе: при большой нагрузке частота и сила дыхательных движений увеличиваются, что приводит к гипервентиляции легких. Что является причиной этого? Усиление тканевого дыхания в мышцах приводит к накоплению молочной кислоты до Ю0...200мг/100мл крови (вместо 15...24 в норме) и недостатку кислорода для окислительных процессов. Такое состояние называется кислородной задолженностью. Молочная кислота, являясь более сильной кислотой, чем угольная, вытесняет из бикарбонатов крови диоксид углерода, в результате этого возникает гиперкапния, что усиливает возбудимость дыхательного центра.
Далее при мышечной работе возбуждаются различные рецепторы: проприорецепторы мышц и сухожилий, механорецепторы легких и воздухоносных путей, хеморецепторы сосудистых рефлексогенных зон, рецепторы сердца и др. От этих и других рецепторов афферентная импульсация также достигает дыхательного центра. При мышечной работе повышается тонус симпатического отдела нервной системы, увеличивается содержание катехоламинов в крови, которые стимулируют дыхательный центр и рефлекторно, и непосредственно. При мышечной работе увеличивается теплопродукция, что также ведет к усилению дыхания (тепловая одышка).
Раздражение различных экстерорецепторов приводит к образованию условных рефлексов. Обстановка, в которой обычно совершается работа (ипподром, ландшафт, взнуздывание, появление наездника, а также время суток), является комплексным стереотипом раздражения, подготавливающим лошадь к последующей работе. Наряду с различными поведенческими актами у животного заранее усиливается работа сердца, повышается артериальное давление, перестраивается дыхание и возникают другие вегетативные изменения.
В начале работы энергия мышцам поставляется за счет анаэробных процессов. В дальнейшем этого оказывается недостаточно и тогда возникает новое стационарное состояние («второе дыхание»), при котором увеличивается вентиляция легких, систолический и минутный объем сердца, кровоток в работающих мышцах.
Таким образом, регуляция дыхания включает два механизма: регуляцию внешнего дыхания, направленную на обеспечение оптимального содержания кислорода и диоксида углерода в крови, т. е. адекватного тканевому метаболизму, и регуляцию кровообращения, создающую наилучшие условия обмена газов между кровью и тканями.
В регуляции вдоха и выдоха большее значение имеют авто-матия дыхательного центра и афферентные импульсы от меха-норецепторов легких и дыхательных мышц, а в регуляции частоты и глубины дыхания - газовый состав крови, цереброспинальной жидкости и афферентные импульсы от хеморецепто-ров кровеносных сосудов, тканей и медуллярных (бульбарных) хеморецепторов.
Дыхание
2. Цель лекции
Проанализировать механизм внешнего дыхания, познакомиться с основными физиологическими показателями легочной вентиляции.
Проанализировать процессы газообмена в легких и тканях, механизмы собственных и сопряженных рефлексов системы дыхания, а также причины его изменения при пониженном и повышенном атмосферном давлении.
З. Задачи лекции. Рассмотреть дыхание на уроне физиологических процессов
Функции дыхательной системы
Типы дыхания
Регуляция дыхания.
Легочные объемы и емкости
Газообмен в легких
Транспорт газов кровью
5. Вопросы для самостоятельной работы,
литература для подготовки
Методические указания к лабораторным занятиям по нормальной физиологии для студентов медицинского института. ПГУ, Пенза 2003.
6. Вопросы для повторения
Анатомия и гистология органов дыхания
Лектор доцент Микуляк Н.И.
Дыхание – это одна из функций организма. Это видно из того, что прекращение дыхания приводит к смерти. Нет дыхания – нет жизни. Почему прекращение дыхания приводит к смерти?
Как вы знаете жизнь это постоянный обмен в-в с окружающей средой. Одним из таких в-в является кислород О 2 , который должен поступать в организм из окружающей среды, а в окружающую среду выбрасывается из организма углекислый газ СО 2 . Кислород необходим организму, т.к. большинство химических реакций в организме являются окислительными с необходимым участием СО 2 . Нет кислорода, нарушаются биохимические процессы, а эти нарушения несовместимы с жизнью. Кроме того, нарушение дыхания ведет к накоплению в организме СО 2 , что губительно сказывается на жизненные отправления в организме. Т.о. дыхание является одной из важнейших функций организма. Нет дыхания – недостаток О 2 – нарушение окислительных биохимических реакций - смерть. Дыхание осуществляется за счет дыхательной системы, т.е. дыхание функция дыхательной системы. Эта ф-ия присуща в некоторой степени коже, слизистой оболочке.
Ф-ция дыхательной системы теснейшим образом взаимосвязана с кровью и ССС. Дыхательная система + кровь + ССС = СКОО (система кислородного обеспечения организма).
Эта взаимосвязь легко выявляется при патологии в организме. Так при воспалении легких, когда нарушается дыхательная ф-ция, наряду с учащением дыхания, усиливается гемодинамика за счет увеличения частоты сокращения сердца, увеличивается кол-во эр., переносчиков О 2 . С другой стороны, нарушения нарушения В ССС, допустим при пороках сердца, когда скорость кругооборота крови уменьшается, усиливается дыхание и гемодинамика.
Дыхание как процесс, складывается из 5 этапов:
1. внешнее дыхание или вентиляция легких или обмен воздуха между внешней средой и альвеолами легких;
2. газообмен (в легких) между альвеолярным воздухом и кровью;
3. транспорт О 2 и СО 2 кровью;
4. обмен газов между кровью и тканями;
5. Тканевое дыхание.
Физиология дыхания изучает первые 4 группы процессов, механизм их регуляции и особенности протекания в различных условиях. Клеточное, т.е. тканевое дыхание изучается в основном биохимией, исследующей тканевые окислительные процессы при которых богатые энергией вещества, содержащиеся в клетке, расщепляются, освобождая скрытую в них энергию.
Вентиляция легких осуществляется за счет периодически сменяющихся вдоха и выдоха.
Рассмотрим вначале инспирацию (механизм вдоха). Вдох – это процесс, обеспечивающий поступление воздуха из окружающей среды в легкие. Вдох – начинается с сокращения дыхательной мускулатуры и мышц диафрагмы. При обычном спокойном вдохе у здоровых людей сокращаются наружние межреберные мышцы и межхрящевые. Это приводит к увеличению размера грудной клетки в сагиттальном направлении и во фронтальном. Почему? В состоянии покоя ребра опущены к низу. При вдохе ребра принимают более горизонтальное положение, поднимаясь кверху. Благодаря чему сечение грудной клетки становиться больше и в поперечном и в продольном направлении. Почему сокращение межреберных мышц ведет не к сближению ребер между собой, а к их поднятию? Это происходит вследствие того, что наружные межреберные мышцы идут от ребра к ребру в косом направлении: сзади и сверху, вперед и вниз. Ребра представляют собой рычаги второго рода в их сочленении с позвоночником. Сила, сокращающейся межреберной мышцы, оказываемая на верхние и низлежащие ребра - одинакова. Но рычаг у низлежащего ребра больше, а следовательно момен6т силы у нижележащего ребра больше, т.е. что легче мышце: поднять нижнее ребро или опустить верхнее или сблизить их? Конечно поднять нижнее ребро. Т.о. подъем ребер ведет к увеличению размеров грудной клетки в сагиттальном и во фронтальном направлении. Кроме того, одновременно сокращается мышца диафрагмы. Это приводит к уплотнению диафрагмы, к опусканию ее купола, вследствие чего увеличивается размер грудной клетки в вертикальном направлении. Опускание диафрагмы на 1 см ведет к увеличению объема на 350 мл. Итак. Грудная клетка увеличивается во всех 3-х направлениях. При спокойном дыхании вдох у мужчин и женщин протекает одинаково. У женщин объем грудной клетки увеличивается за счет преимущественного сокращения межреберных мышц. Это так называемый, грудной тип дыхания или реберный. Такой же тип дыхания у кошек. У мужчин объем грудной клетки увеличивается преимущественно за счет диафрагмы. Это так называемый брюшной или диафрагмальный тип дыхания. Такое дыхание у кроликов.
Тип дыхания не является постоянным и зависит от вида выполняемой работы. Так при переносе груза, дыхание осуществляется за счет движения диафрагмы. При усиленном дыхании (при одышке) в акте вдоха участвуют ряд дополнительных вспомогательных мышц: стерноклейдомастоидей, леватор скапуле, пекторалис майор и минор и т.д.
Итак. Вдох начинается с сокращения дыхательной мускулатуры, что приводит к увеличению объема легких. Легкие всегда следуют за грудной клеткой. Почему? Остановимся на этом.
1. Это обусловлено герметичностью грудной клетки:
2. Свойствами легочной ткани.
Для того, чтобы понять этот процесс надо вспомнить о так называемой модели Дондерса: берут стекл. бутыль с резиновым дном, верхнее отверстие бутыли закрыто пробкой через которую пропущена стеклянная трубка, на которую надевается трахея с легкими от мелкого животного (крысы или кролика). Сбоку в бутыль вмонтирован манометр. На легкие изнутри, т.е. через стекл. трубку действует давление в 1 атм. Снаружи, т.е. из бутыли на поверхность легких также действует давление = 1 атм. Две силы равны, легкие находятся в состоянии покоя. Если оттянуть резиновое дно, то давление в бутыли понизиться, возникает разность давления, действующих на легкие на внутреннюю и наружную стороны. Через трубку действует давление больше. Поэтолму воздух (наружний) поступает в легкие и они растягиваются. Одновременно с этим следует отметить. Что давление в бутыли остается меньше атмосферного.
Закон Бойля-Мариотта р1/р2=v1/v2 или р1v1=р2v2
А теперь перенесемся от этой модели к целому организму.
Легкие покрыты висцеральным листком плевры. Внутренняя поверхность грудной клетки покрыта париетальным листком плевры. Между ними имеется плевральная полость (щель). Между ними имеется некоторое количество жидкости, которая обеспечивает смазку листков, необходимую для уменьшения трения между ними. Плевральная полость герметично замкнута. У человека две плевральные полости. Если человеку ввести в плевральную полость иглу, соединенную с манометром, то мы увидим, что там давление ниже атмосферного на несколько мм. В состоянии свободного выдоха оно = 7 мм.рт.ст. При вдохе оно становиться = 9-10 мм.рт.ст. При максимальном выдохе = 2-3 мм.рт.ст. При максимальном вдохе до 30 мм.рт.ст. А если закрыть дыхательные пути и сделать попытку вдоха (опыт Мюллера), то оно становиться ниже атмосферного на 50-50 мм.рт.ст. Это давление называется отрицательным давлением. Отрицательное давление – это разность между атмосферным давлением и давлением в плевральной полости. Чем же обусловлено отрицательное давление?
Это обусловлено свойствами легочной ткани.
1. растяжимость
2. эластичность.
Если зажать трахею у мертвого животного, вскрыть грудную клетку, то мы увидим, что легкие занимают всю грудную клетку, т.е. они находятся в растянутом состоянии. Если через трахею подать воздух под давлением, то легкие растянуться еще больше. Т.е. легочной ткани присуща растяжимость. Это свойство для легочной ткани присуще в большей степени, чем для любой другой.
Если открыть трахею, продолжая опыт. То из легких выбрасывается воздух и легкие уменьшаются в размере. Это связано с эластичностью легочной ткани.
Эластичность – это способность ткани принимать первоначальный объем или форму. И обусловлена она содержанием большого количества эластических волокон. За счет этих волокон создается эластическая тяга легких – которая постоянно существует в организме, т.к. легкие всегда находятся в растянутом состоянии. Это обусловлено тем. Что грудная клетка
1. имеет больший объем, чем легкие, и она
2. быстрее растет, чем легкое.
Эластическая тяга легких старается постоянно привести объем легких к минимальному, т.е. оторвать висцеральный от париетального. Но т.к. плевральная полость герметически замкнута, то в этой полости создается несколько разреженное пространство, т.е. отрицательное давление.
Эластическая тяга легких зависит:
1. от наличия в стенке альвеол большого кол-ва эластических волокон,
2. обусловлена поверхностным натяжением стенки альвеол.
Что будет с легкими, если нарушить герметичность плевральной щели? Давление на наружную и внутреннюю поверхность легких в этом случае = атмосферному. Но остается эластическая тяга легких, за счет которой легкие сжимаются, принимая минимальный объем. Это состояние называется пневмотораксом. При этом легкие спадаются и дыхательная ф-ция выключается. Пневмоторакс может быть односторонним. Пневмоторакс иногда применяется для лечения.
Т.о. механизм вдоха слагается из:
1. сокращение межреберных мышц и мышц диафрагмы
2. увеличение размеров грудной клетки
3. увеличение объема легких
4. понижение давления в легких
5. поступление воздуха в легкие
Выдох – пассивный (спокойный). Происходит под влиянием силы тяжести грудной клетки и давления органов брюшной полости. Но может быть и активным – форсированным, когда к перечисленным силам уменьшающим объем, присоединяется сокращение внутренних межреберных косых мышц, задних внутренних зубчатых мышц и мышц живота.
Дыхательная мускулатура, обеспечивающая вдох, совершает большую работу. Эта работа необходима для преодоления сопротивления, которое складывается из статического и динамического.
Статическое давление (эластическое) включает
1. вес грудной клетки, которую надо поднять
2. сопротивление сжатию органов брюшной полости, которые оттесняются опускающейся диафрагмой.
3. К статическому сопротивлению относится также преодоление эластического сопротивления ткани легкого при ее растяжении.
При глубоком дыхании статическое сопротивление возрастает.
Динамическое сопротивление (вязкостное или неэластическое) различают
1. тканевое сопротивление
2. воздушное сопротивление
К тканевому сопротивлению относится:
1. трение между листками плевры
2. трение между сердцем и легкими
Воздушное сопротивление, оказываемое со стороны дыхательных путей движущемуся воздуху, это сопротивление зависит от:
1. длины дыхательных путей
2. их диаметра
3. характера движения воздушной струи
4. скорости движения воздуха.
Может ли меняться длина дыхательных путей? Может. Длина дыхательных путей меняется в зависимости от того, дышит человек через нос или через рот. В первом случае длина больше, а значит и воздушное сопротивление возрастает. Длина дыхательных путей возрастает во время вдоха, при выдохе уменьшается. Значительно увеличивается длина дыхательных путей в противогазах. Для того чтобы уменьшить сопротивление и уменьшить работу дыхательной мускулатуры бегуны на короткие дистанции дышат через рот. Но постоянное дыхание через рот чревато большими опасностями. Во-первых, часто возникают простудные заболевания верхних дыхательных путей. Во-вторых, отсутствие дыхания через нос ведет к снижению умственных способностей – к слабоумию. В-третьих, - нарушается вентиляция легких (струя воздуха идущая через нос раздражает рецепторы слизистой носа – импульс в дыхательный центр – усиление дыхания). В-четвертых, выключение носового дыхания ведет к нарушению половой потенции. Это возникает при полипозе носа, когда разрастается лимфатическая ткань в носу.
Воздушное сопротивление зависит от диаметра воздухоносных путей. Диаметр дыхательных путей постоянен у здоровых людей. Он несколько увеличивается при вдохе и уменьшается при выдохе, поэтому выдох происходит медленнее. Чем вдох на 5 – 10 %. Диаметр дыхательных путей уменьшается у курящих людей. К старости при различных заболеваниях органов дыхания (при бронхиальной астме, когда резко уменьшается диаметр, особенно при выдохе, поэтому у этих больных резко затруднен выдох).
Воздушное сопротивление зависит от характера движения воздушной струи. Различают 2 типа движения воздуха: ламинарное и турбулентное.
Ламинарный тип – когда все слои воздуха движуться параллельно – сопротивление наименьшее. Воздух при этом движется клиновидным фронтом. Этот тип дыхания возможен при гладких стенках воздухоносных путей и при относительно небольшой скорости воздуха, а это может быть только при спокойном дыхании.
Турбулентный тип (вихревой), когда частицы воздуха постоянно перемешиваются между собой и сопротивление резко возрастает. Это наблюдается при частом дыхании, при различных заболеваниях, когда нарушается гладкая поверхность дыхательных путей.
Воздушное сопротивление зависит от скорости движения воздуха. При этом больше динамическое сопротивление. В свою очередь скорость движения воздуха зависит от диаметра дыхательных путей т от интенсивности дыхания.
Между статическим и динамическим сопротивлением существует зависимость, которая определяется частотой дыхания. При частом дыхании увеличивается динамическое сопротивление, а при редком – статическое. Мингимальное сопротивление имеет место при частоте дыхания 15 раз в 1 мин. и называется эйпное. Если дыхание редкое (называется брадипное, частое – тахипное).
Легочные объемы и емкости .
Для суждения о легочной вентиляции, т.е. о внешнем дыхании используют определение легочных объемов и емкостей. По цифрам этих показателей можно составить представление о внешнем дыхании. Это используют чаще при определении физического развития человека.
Легочные объемы:
1. ДО – дыхательный объем – это кол-во воздуха, которое поступает и выделяется при спокойном дыхании. ДО=500 мл. (300-900)
2. РОВд – резервный объем вдоха – это кол-во воздуха, которое можно вдохнуть после спокойного вдоха. РОВд=1500 мл. (1,5 – 1,8)
3. РОВыд - резервный объем выдоха – это кол-во воздуха, которое можно выдохнуть после обычного выдоха. РОВд=1500 мл.
4. ОО – остаточный объем – это воздух, который остается после максимального выдоха. Можно определить при вскрытии. ОО = 1500 мл. (1,0 – 1,5)
5. КО – коллапсный объкм. Остается после спадения легких, после выдоха остаточного объема. Поэтому легкие человека, хотябы раз вдохнувшего воздух не тонут в воде. Это используется в судебно-медицинской практике. КО = 150 мл.
Легочные емкости:
Кроме легочных объемов выделяют легочные емкости, которые представляют 2 или несколько объемов, измеренных суммарно:
1. ОЕЛ – общая емкость легких = 5150. ОЕЛ=ДО+РОВд+РОВыд+ОО+КО
Метод плетизмографии или метод газовой ди…..
2. ЖЕЛ – жизненная емкость легких. Это кол-во в-ха, которое можно выдохнуть после максимального вдоха. ЖЕЛ=ДО+РОВд+РОВыд=3500 мл.
(3,5- 5,0) муж, (3,0 – 4,0) жен.
3. ЕМ. Выд – емкость максимального выдоха – воздух, который можно выдохнуть при максимальном выдохе, после спокойного вдоха. ЕМВыд=ДО+РОВыд=2000мл. (2,0-2,3)
4. ЕМВд - емкость максимального вдоха. ЕМВд=ДО+РОВд =2000мл
5. ФОЕ – функциональная остаточная емкость легких – кол-во в-ха, которое остается в легких после спокойного выдоха. ФОЕ=ОО+РОВыд=3000мл.
Функциональные показатели и пробы дыхания.
Легочные объемы и емкости дают право приблизительно представить состояние дыхательного аппарата. Более подробно и точнее о состоянии дыхательного аппарата можно судить изучая функциональные показатели легких и давая различную нагрузку на легкие.
Существует очень много показателей, но более часто применяются в клинике следующие:
1. ЧД – частота дыхания. Средняя 14 – 15 в 1 мин, варьирует от 20 до 40. Если реже или чаще, то это уже нарушение.
2. ГД – глубина дыхания – кол-во в-ха. Которое забирается легкими при вдохе.
3. МОД – минутный объем дыхания – кол-во в-ха, которое проходит через легкие при обычном дыхании: МОД=ЧД*ГД/ДО/=16*500=8000мл.
Минутный объем у здоровых людей колеблется от 6 до 8 л. МОД зависит от возраста, пола и роста, от веса тела. Поэтому при определении МОД, его сравнивают с должным минутным объемом дыхания ДМОД.
ДМОД – определяется по нормограмме и по эмпирическим выведенным формулам:
ДМОД (у мужчин)= 3,2*5 м 2 (поверхность тела)
ДМОД (у женщин)= 3,7*5 м 2 (поверхность тела)
4. МВЛ – кол-во воздухха, которое проходит через легкие в 1 мин. при дыхании max глубоком и max частом.
(130-140л/мин у муж., 110 – 120 л/мин у жен.)
5. РД – это разность между МВЛ и МОД
РД=МВЛ-МОД=120 – 130 л
6. ЖПЛ – это отношение ЖЕЛ к массе тела.
ЖЕЛ/М= 75 мл/кг у муж, и 65 мл/кг у жен.
7. Максимальная скорость движения воздуха – МСДВвд = 3,2 м/с
МСДВвыд=2,8 м/с
8. АВЛ – это показатель того количества воздуха. Который участвует непосредственно в газообмене. Часть воздуха не участвует в газообмене, это та часть. Которая находиться в носовой полости гортани, бронхов. Бронхиолах. Эти дыхательные пути называются мертвым пространством, его объем равен 150 мл. АВЛ=(ДО-ОМП)*ЧД=350*16=5,6 л
МОД= 9 МОД= 9
1) ЧД= 30 2) ЧД= 15
ГД= 300 мл ГД = 600мл
АВЛ!=150*30=4,5л АВЛ!=450*15=6,75л
Альвеолярное дыхание зависит от частоты и глубины. Мертвое пространство играет роль:
1. буфера между альвеолярным и атмосферным воздухом. При каждом вдохе последние порции воздуха задерживаются в мертвом пространстве, поэтому альвеолярный воздух мало меняет свой состав. К концу выдоха в альвеолах находиться ФОЕ функциональная остаточная емкость.
При вдохе альвеолярный воздух обновляется не весь, а всего 1/9 часть. (3150+ 350)
2. роль механического фильтра. Вдыхаемый воздух соприкасается со слизистой оболочкой и очищается.
3. выдыхаемый воздух увлажняется
4. Роль температурного реле. Обеспечивается дыхание при резких перепадах температуры.
В легкие при вдохе поступает атмосферный воздух. Его состав:
О 2 – 21%, СО 2 – 0,63%, N 2 – 79%.
Атмосферный воздух проходя по дыхательным путям смешивается с воздухом альвеол, поэтому в альвеолярном воздухе содержиться:
О 2 – 14%, СО 2 – 5,5%, N 2 – 79%.
Состав альвеолярного воздуха постоянен.
При выдохе альвеолярный воздух смешивается с воздухом мертвого пространства, поэтому О 2 – 16%, СО 2 – 4,5%, N 2 – 79%. Основное назначение вентиляции легких – обеспечить постоянство состава альвеолярного воздуха.
Газообмен в легких.
Обмен газа между альвеолярным воздухом и кровью происходит в альвеолах. Легочная ткань и кровь разделяет альвеолярно-капиллярный барьер, который образован 2-мя слоями клеток – слой эндотелия и слой эпителия, толщина 0,5 мкм. За 1 сек через барьер проходит СО 2 и О 2 , за 1 сек уравнивается состав альвеолярного воздуха и крови. Барьер имеет высокую проницаемость для газов.
Имеется большое количество альвеол, их кол-во в одном легком 300 – 400 млн., общая площадь = 80 – 100 м 2 . Через альвеолярную поверхность за 1 мин. в организм поступает в кровь 250 мл О 2 и удаляется 250 мл СО 2 .
Необходимо для этого при МОК – 5 л. крови (малый круг).
Для газообмена имеет значение парциальное давление. И напряжение газов.
Парциальное давление – это то давление, которое приходиться на долю газа в смеси, если газ находиться в жидкости, то давление газа в жидкости называется напряжением.
Парциальное давление в альвеолярном воздухе: 760-50=710 мм.рт.ст.
Р О2 = 710*14/100= 100 мм.рт.ст.
Р СО2 = 710*5,5/100=40 мм.рт.ст
Р N 2 = 575 мм.рт.ст.
Напряжение газов в венозной крови: О 2 – 40, СО 2 – 46
В артериальной крови: О 2 – 100, СО 2 – 60
В тканях О 2 – 0, СО 2 – 60
Диффузия газов осуществляется при наличии разности между парциальным давлением и напряжением газов.
Газы будут диффундировать в сторону меньшего давления. В альвеолах легкого в венозную кровь идет О 2 , а СО 2 идет при разности градиента давления равен 6 . этот градиент достаточен для того, чтобы удалить из организма 200 мл СО 2 .
Проницаемость барьера не одинакова для всех газов. Для О 2 составляет 25 мл в мин, т.е ч/з барьер в мин. может пройти 25*60=1500мл О 2 .
В норме же = 250 мл.
Газообмен легких осуществляется за счет разности парциального давления газов в альвеолярном воздухе и напряжением их в венозной крови. Обмену газов содействует высокая проницаемость барьера для газов.
Назначение газообмена для газов – ограничение поступления О 2 в кровь и удаление СО 2 . Интенсивность обмена составляет в среднем 250 мл О 2 , 200 мл СО 2 /мин.
Транспорт газов кровью.
В 100 мл артериальной крови О 2 =20 мл. СО 2 = 52 мл.
В 100 мл венозной крови будет находиться О 2 =12 мл. СО 2 = 58 мл.
Часть газов в крови будет находиться в физически растворенном состоянии.
В 100 мл крови растворяется 0,3 мл О 2 1 мл N 2 и 2-3 мл. СО 2 . Основная часть газов находиться в связанном состоянии.
Переносчиком O2 от легких к тканям и С02 от тканей в легкие является кровь. В растворенном состоянии в плазме крови дыхательных газов переносится мало. 6 основном они транспортируются в связанном состоянии.
Транспортировку кислорода. Количество растворенного в крови кислорода составляет 0,03 мл на 1 л крови / мм рт. ст. В артериальной крови Ро2 = 100 мм рт. ст., поэтому в 1 л ее транспортируется растворенного кислорода только 3 мл.
Главная часть O2 транспортируется кровью в виде химического соединения с гемоглобином - оксигемоглобин: Нb + O2 → НbO2. В связи с тем, что гемоглобин состоит из 4 субъединиц, 1 моль гемоглобина может связать 4 моли О2. Это означает, что 1 г Нb присоединяет 1,34 мл O2 (число Хюфнер). Зная число Хюфнер (1,34) и количество гемоглобина ВИЧ крови (150 г), можно рассчитать кислородную емкость крови (КЕК), которая составит: 1,34 мл O2 150 = 200 мл О2 / л. КЕК - это максимальное количество O2, который может связать кровь при полном насыщении гемоглобина кислородом.
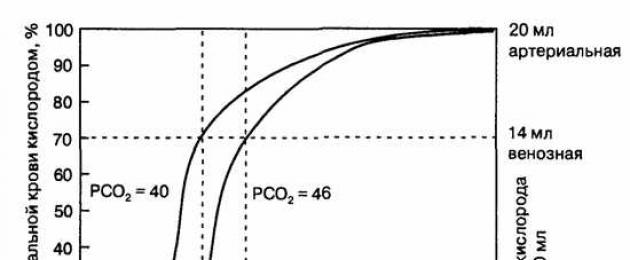
Однако в естественных условиях гемоглобин оксигенуеться не полностью. Согласно закону действующих масс, насыщения гемоглобина кислородом (в%) зависит от напряжения кислорода (Ро2) в крови. Графически эту зависимость отражает так называемая кривая диссоциации оксигемоглобина, что S-образную форму (рис. 11.14). Комбинация первого гема в молекуле Нb с O2 усиливает сродство второго тему для O2, а оксигенация другой - усиливает сродство третьего и так далее, следовательно, родство Нb для четвертой молекулы O2 в несколько раз больше, чем для первой.
Когда напряжение O2 равна 0, в крови находится восстановленный гемоглобин - дезоксигемоглобин. Повышение напряжения O2 сопровождается увеличением количества оксигемоглобина, но этот прирост имеет не линейный характер, а S-образный. Особенно быстро нарастает концентрация оксигемоглобина при повышении Ро2 от 10 до 40 мм рт. ст. При Ро2 равном 60 мм рт. ст. оксигемоглобин достигает 90%. При дальнейшем нарастании Ро2 насыщения гемоглобина крови идет очень медленно и кривая диссоциации переходит из крутого к пологому роста.
Пологая часть свидетельствует, что в этих условиях содержание оксигемоглобина мало зависит от напряжения кислорода (Ро2) и его парциального давления во вдыхаемом и альвеолярном воздухе. Например, при поднятии на высоту 2 км атмосферное давление снижается с 760 до 600 мм рт. ст., а содержание оксигемоглобина - на 3%. Таким образом, пологая часть кривой диссоциации отражает способность связывать большее количество 02, несмотря на умеренное снижение парциального давления во вдыхаемом воздухе.
Крутой наклон среднего участка кривой диссоциации оксигемоглобина свидетельствует о благоприятной ситуации для отдачи кислорода тканям. При необходимости органов в кислороде он должен высвобождаться в достаточном количестве даже в условиях отсутствия значительных сдвигов Ро2 в артериальной крови. В состоянии покоя Ро2 в области венозного конца капилляра составляет 40 мм рт. ст. (5,3 кПа), что соответствует 73 % насыщения (см. Рис. 11.14).
РИС. 11.14. Кривая диссоциации оксигемоглобина в норме - красная линия, и под влиянием родственных кислорода факторов - коричневая и синяя. Стрелочки вверх - увеличение величины действующих факторов, вниз - их уменьшения. Факторами, влияющими на сродство гемоглобина к кислороду, являются: pH, Рсо2, температура, концентрация 2,3-дифосфоглицерата (2,3-ДФГ), HbF
Смещение кривой диссоциации оксигемоглобина вправо. Уменьшение pH, увеличение Рсо2, повышение температуры крови, увеличение концентрации в эритроцитах 2,3-ДФГ сдвигает кривую справа - это значит, что при том же Ро2 диссоциация оксигемоглобина увеличивается: НЬ + 02 "- НЬ02. Это имеет место в капиллярах, которые приносят кровь к мышцам во время физической работы, когда снижается pH, повышается Рсо2, растет температура крови.
Эритроциты богаты 2,3-ДФГ, который является продуктом гликолиза. Это високообминний анион, который присоединяется к β-цепей дезоксигемоглобином. Один моль дезоксигемоглобином связывает 1 моль 2,3-ДФГ: НbO2 + 2.3-ДФГ → Нb-2,3-ДФГ + O2.
К факторам, которые влияют на концентрацию 2.3-ДФГ в эритроцитах, принадлежит pH. Поскольку ацидоз ингибирует гликолиз в эритроцитах, то при снижении pH концентрация 2,3-ДФГ уменьшается. При гипоксии увеличивается интенсивность гликолиза и соответственно - концентрация 2,3-ДФГ, что способствует увеличению диссоциации оксигемоглобина. Такие изменения имеют место при адаптации человека к гипоксии в условиях высокогорной местности.
Гемоглобин плода - фетальный гемоглобин (HbF) имеет большее сродство к кислороду, чем гемоглобин взрослых - НbА, что облегчает поступление кислорода от матери к плоду.
Смещение кривой диссоциации оксигемоглобина влево. При росте pH, снижении Рсо2, температуры и концентрации 2,3-ДФГ кривая диссоциации оксигемоглобина смещается влево; это означает, что меньше кислорода диффундирует в клетки организма.
Транспортировка СO7. Углекислый газ транспортируется кровью в виде:
■ физически растворенного (СО2) и в форме угольной кислоты (Н7С03) - 12%;
■ карбаминовой соединения (NHCOOH), упрощенно - карбогемоглобин 11%;
■ бикарбонатных ионов в эритроцитах - 27%;
■ остальные - 50% растворенные в плазме в форме НСО3-.
Напряжение СО2 в артериальной крови, поступающей в тканевые капилляры, составляет 40 мм рт. ст. В клетках вследствие метаболизма образуется значительное количество СО2 (Рсо2 - 70 мм рт. Ст.), Который благодаря градиенту напряжения диффундирует в плазму крови и в эритроциты.
В эритроцитах большая часть СО2 связывается с водой и под воздействием фермента карбоангидразы образуется угольная кислота СО2 + Н2О = Н2СO3, которая распадается на ион водорода и бикарбонат Н2СO3 → H + + НСО 3. НСO3 переходит в плазму в обмен на анион хлора (хлоридное сдвиг) (рис. 11.15). Ион Н + связывается с восстановленным НЬ (дезоксигемоглобином), образуя слабую кислоту ННb, которая соединяется с СО2.
В то же время кислород диффундирует в клетки организма, способствует диффузии СО2 в кровь, потому дезоксигемоглобин является слабой кислотой, чем оксигемоглобин, и может присоединять больше ионов водорода, в результате чего повышается степень диссоциации Н2СO3 → Н + + НСО 3, увеличивается транспортировки СО2 кровь "ю (эффект Холдейна).
В плазме НСО3- взаимодействует с катионами и создает соли угольной кислоты (NaHCO3), которые транспортируются в легочных капилляров.
Небольшая часть СО2 в эритроцитах соединяется с конечными аминогруппами глобиновой части молекулы гемоглобина, образуя соединение карбамата: HbNH2 + СО2 = HbNHCOOH + Н +. Кроме этого, карбаминови соединения в малом количестве образуются с белками плазмы крови.
В легких все реакции идут наоборот. НСО3- входит в эритроциты в обмен на ионы Сl - . Ионы Н + вместе с ионами НСО3- образуют Н2СO3, которая расщепляется под воздействием карбоангидразы на СО2 и Н2О. Углекислый газ диффундирует в альвеолы и выдыхается.