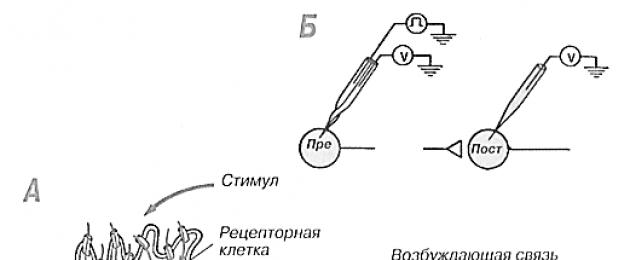
Понятие о сенсорных рецепторах. Основным компонентом периферического отдела сенсорных систем является рецептор . Он представляет собой высокоспециализированную структуру (у первичночувствующих рецепторов - это видоизмененный дендрит афферентного нейрона, у вторичночувствующих - это сенсорная рецепторная клетка), которая способна воспринимать действие адекватного раздражителя внешней или внутренней среды и трансформировать его энергию в конечном итоге в потенциалы действия - специфическую активность нервной системы. Здесь следует напомнить, что понятие «рецептор» (от лат. гесерio, гесерtum - брать, принимать) в физиологии используется в двухзначениях. Во-первых, для обозначения специфических белков клеточной мембраны или цитозоля, которые предназначены для детекции гормонов, медиаторов и других биологически активных веществ. Такие рецепторы принято называть мембранными, клеточными, или гормональными (например, альфа-адренорецепторы). Во-вторых, для обозначения рецепторов как компонентов сенсорной системы. Эти рецепторы часто называют сенсорными рецепторами, или сенсорными рецепторными клетками.
Классификация рецепторов. В зависимости от того, из внутренней или внешней среды воспринимаются раздражения, все сенсорные рецепторы подразделяют на экстероцепторы и интерорецепторы . Экстерорецепторы воспринимают сигналы внешней среды. К ним относят фоторецепторы сетчатки глаза, фонорецепторы кортиевого органа, вестибулорецепторы полукружных каналов и мешочков преддверия, тактильные, температурные и болевые рецепторы кожи и слизистых оболочек, вкусовые рецепторы языка, обонятельные рецепторы носа. Среди интерорецепторов различают висцерорецепторы, предназначенные для детекции изменений внутренней среды, и проприрецепторы (рецепторы мышц и суставов, т.е. опорно-двигательного аппарата). Висцерорецепторы представляют собой различные хемо-, механо-, термо-, барорецепторы внутренних органов и кровеносных сосудов, а также ноцицепторы.
По характеру контакта со средой экстерорецепторы делятся на дистантные , получающие информацию на расстоянии от источника раздражения (зрительные, слуховые и обонятельные) и контактные - возбуждающиеся при непосредственном соприкосновении с раздражителем (вкусовые, тактильные).
В зависимости от вида модальности воспринимаемого раздражителя, т.е. от природы раздражителя, на который рецепторы оптимально настроены, сенсорные рецепторы делятся на 6 основных групп : механорецепторы, терморецепторы, хеморецепторы, фонорецепторы, ноцицепторы и электрорецепторы (последние выявлены только у некоторых рыб и амфибий).
Механорецепторы приспособлены к восприятию механической энергии раздражающего стимула. Они входят в состав соматической (тактильной), скелетно-мышечной, слуховой, вестибулярной и висцеральной сенсорных систем, а также (у рыб и амфибий) сенсорной системы боковой линии. Терморецепторы воспринимают температурные раздражения, т.е. интенсивность движения молекул, и входят в состав температурной сенсорной системы. Они представлены тепловыми и холодовыми рецепторами кожи, внутренних органов и термочувствительными нейронами гипоталамуса. Хеморецепторы чувствительны к действию различных химических веществ и входят в состав вкусовой, обонятельной и висцеральной сенсорных систем. Фоторецепторы воспринимают световую энергию и составляют основу зрительной сенсорной системы. Болевые (ноцицептивные) рецепторы воспринимают болевые раздражения, в том числе механоноцицепторы - действие чрезмерных механических стимулов, хемоноцицепторы - действие специфических болевых медиаторов; они являются начальным компонентом ноцицептивной сенсорной системы. Электрорецепторы, выявленные в составе боковой линии ряда рыб и амфибий, чувствительны к действию электромагнитных колебаний.
Следует подчеркнуть, что в процессе эволюции отбирались те рецепторы и соответствующие им сенсорные системы, которые обеспечивали каждый организм достаточным количеством информации, необходимой для его нормального существования и адаптации во внешней среде. В этом плане можно привести образно сказанную фразу (А.Д. Ноздрачев и соавт., 1991): «У человека не обнаружены электрорецепторы, существующие у рыб; нет рецепторов, воспринимающих прямое инфракрасное излучение, как у гремучей змеи; глаз человека не воспринимает поляризацию света, как глаза некоторых насекомых, его ухо не ощущает ультразвуковых колебаний, как слуховой аппарат летучих мышей и многих ночных млекопитающих». Но, в целом, имеющиеся у человека сенсорные системы позволяют ему успешнее других представителей животного мира осваивать Землю.
Кроме представленных двух классификаций важным является деление всех сенсорных рецепторов в зависимости от их структуры и взаимоотношения с афферентным сенсорным нейроном на два больших класса - первичночувствующие (первичные) и вторичночувствующие (вторичные) рецепторы . Это определяет избирательную чувствительность рецептора к адекватным раздражителям (у вторичночувствующих она намного больше, чем у первичночувствующих), а также последовательность трансформации энергии внешнего сигнала в потенциал действия нейрона.
К первичным сенсорным рецепторам относят те рецепторы, которые представляют собой видоизмененное, специализированное окончание дендрита афферентного нейрона. Это означает, что афферентный нейрон непосредственно (т.е. первично) взаимодействует с внешним стимулом. К первичночувствующим рецепторам относятся отдельные виды механорецепторов (свободные нервные окончания кожи и внутренних органов), холодовые и тепловые терморецепторы, ноцицепторы, мышечные веретена, сухожильные рецепторы, суставные рецепторы, обонятельные рецепторы.
Вторичные рецепторы - это специально приспособленные для восприятия внешнего сигнала клетки ненервного происхождения, которые при своем возбуждении в ответ на действие адекватного раздражителя передают сигнал (как правило, с выделением медиатора из синапса) на дендрит афферентного нейрона. Следовательно, в этом случае нейрон воспринимает раздражитель косвенно, опосредовано (вторично) за счет возбуждения сенсорной клетки-рецептора (рецептирующей клетки). К вторичночувствующим рецепторам относятся многие виды механорецепторов кожи (например, тельца Пачини, диски Меркеля, клетки Мейсснера), фоторецепторы, фонорецепторы, вестибулорецепторы, вкусовые рецепторы, а также электрорецепторы рыб и амфибий.
Адаптация сенсорных рецепторов. Сенсорные рецепторы способны к адаптации, которая состоит в том, что при постоянном воздействии стимула на сенсорный рецептор его возбуждение слабеет, т.е. снижается величина рецепторного потенциала, а также частота генерации потенциалов действия афферентным нейроном. Подобное явление наблюдается и при гормонрецепторном взаимодействии. В этом случае оно называется десенситизация и связано с нарушениями передачи сигнала «вниз по течению». Адаптация сенсорных рецепторов имеет еще более сложную природу. С одной стороны, она зависит от процессов, которые происходят на этапе взаимодействия сенсорного стимула с «активным центром» сенсорного рецептора (по сути, это явление десенситизации). С другой стороны, адаптация рецепторов связана с потоком импульсов, приходящим к сенсорному рецептору по эфферентным волокнам от вышележащих нейронов мозга (в том числе от нейронов ретикулярной формации), т.е. является активным процессом. В определенной степени, адаптация может быть обусловлена свойствами и состоянием вспомогательных структур периферического отела сенсорной системы. В целом, адаптация проявляется в снижении абсолютной и повышении дифференциальной чувствительности сенсорной системы. Скорость адаптации для разных рецептов различна: наибольшая для тактильных рецепторов, а наименьшая – для вестибулярных и проприорецепторов. Благодаря высокой скорости адаптации тактильных рецепторов мы быстро перестаем ощущать надетые очки, часы или одежду, а благодаря низкой скорости адаптации мышечных рецепторов можем совершать высоко координированные и четкие движения.
Основные этапы преобразования энергии внешнего стимула в рецепторный потенциал (механизмы возбуждения сенсорных рецепторов). При всем многообразии морфофункциональных особенностей сенсорных рецепторов общую схему этого процесса можно представить в виде некоторой обобщенной схемы. В первичных рецепторах условно можно выделить пять основных этапов трансдукции сенсорного сигнала: 1) взаимодействие воспринимаемого стимула с «активным» участком сенсорного рецептора; 2) изменение ионной проницаемости мембраны; 3) уменьшение уровня мембранного потенциала сенсорно го рецептора, т.е. генерация рецепторного потенциала, уровень которого зависит от величины воспринимаемого стимула; 4) генерация потенциалов действия или увеличение частоты генерации спонтанных потенциалов действия в соме афферентного нейрона (аксонном холмике); 5) распространение потенциалов действия по аксону до второго афферентного нейрона данной сенсорной системы. Во вторичночувствующих сенсорных клетках первые три этапа идут по такой же схеме; затем добавляется еще два промежуточных этапа - 4а) выделение под влиянием рецепторного потенциала квантов медиатора (например, ацетилхолина) в синапсе рецепторной клетки; 5а) ответ дендрита афферентного нейрона на выделение медиатора генерацией возбуждающего постсинаптического потенциала, или генераторного потенциала. Остальные два этапа (4 и 5) идут так же, как и в первичночувствующих рецепторах. Единственным исключением из этого правила является цепь событий в зрительной сенсорной системе, в которой в ответ на действие света фоторецепторная клетка повышает свой мембранный потенциал, в результате чего в ней снижается продукция тормозного медиатора, что приводит к конечном итоге к возбуждению биполярного нейрона, который в свою очередь возбуждает ганглиозную клетку.
Рецептор - специальный орган или клетка, воспринимающие раздражение.
Рецепторы - лат. recipere, получать - концевые образования афферентных нервных волокон , воспринимающих раздражения из внешней или из внутренней среды организма и преобразующие физическую или химическую энергию раздражителей в возбуждение (нервные импульсы), передаваемое по чувствительным нервным волокнам в ЦНС .
Сенсорные рецепторы - это специализированные нейроны , которые действуют как преобразователи энергии окружающей среды. Некоторые из них (например, фоторецепторные клетки глаза) снабжают организм данными о внешней среде, другие (например, барорецепторы , реагирующие на изменения давления крови в артериях) сообщают сведения о внутренней среде. Эта информация, закодированная в виде последовательностей импульсов, затем передается в ЦНС первичными афферентными нейронами . Клеточные тела первичных афферентных нейронов находятся в ганглиях задних (дорсальных) спинномозговых корешков или черепных нервов . Периферический отросток каждого первичного афферентного нейрона направляется в составе периферического нерва дистально к соответствующему сенсорному рецептору, а центральный отросток поступает в ЦНС через задний корешок либо черепной нерв ( рис. 33.1 , а).
Рецептор воспринимают внешнее раздражение и трансформируют его энергию в энергию нервного импульса. В зависимости от локализации различают несколько типов рецепторов: 1) экстерорецепторы , воспринимающие раздражения внешней среды, расположены в коже, слизистых оболочках и органах чувств; 2) интерорецепторы , получающие раздражение, главным образом, при изменениях химического состава внутренней среды и давления, расположены в сосудах, тканях и органах; 3) проприорецепторы заложены в мышцах, сухожилиях, связках, фасциях, надкостнице, суставных капсулах.
Каждая рецепторная клетка воспринимает раздражения с определенной зоны - рецептивного поля , представляющего собой все точки периферического отдела анализатора, возбуждение которых влияет на данный нейрон. Чем больше число рецепторов, воспринимающих стимул, и частота нервных импульсов, тем больше размеры воспринимаемого раздражения и его сила.
Простейший рецептор состоит из одной неспециализированной нервной клетки - первичного сенсорного нейрона , периферическое окончание которого способно воспринимать раздражение и порождать нервный импульс, направляющийся в центральную нервную систему , например, кожные механорецепторы типа телец Пачини . Более сложные рецепторы называются вторичными сенсорными клетками и представляют собой видоизмененные эпителиальные клетки, способные воспринимать стимулы. Эти клетки образуют синапсы с соответствующими сенсорными нейронами , передающими импульсы в мозг , таковы, например, клетки вкусовых луковиц .
Наиболее сложными рецепторами являются органы чувств , которые состоят из большого числа чувствительных клеток и связанных с ними вспомогательных приспособлений. Вспомогательные структуры часто выполняют две функции - препятствуют действию посторонних стимулов и усиливают действие нужного стимула.
Все рецепторы преобразуют энергию стимула в локальный нераспространяющийся электрический сигнал, который возбуждает нервные импульсы в сенсорном нейроне. Многообразные стимулы, закодированные в виде нервных импульсов, передаются в мозг, где они расшифровываются и используются для выработки нужной ответной реакции. Характер ответа, его величина и продолжительность находятся в прямой зависимости от природы стимула.
А. Понятия. Различают сенсорные и эффекторные рецепторы. Сенсорные рецепторы воспринимают изменения внешней или внутренней среды организма и обеспечивают поступление информации в ЦНС. Эффекторные рецепторы рабочих органов - эффекторов (сердце, скелетная мышца, желудок и т.д.) воспринимают сигналы (команды) от нервной системы в виде нервных импульсов, они реагируют также на гуморальные вещества, циркулирующие в крови (гормоны, медиаторы, метаболиты). Нервные клетки передают сигналы друг другу также посредством рецепторов, которые тоже являются эффекторными. В данном разделе рассматриваются только сенсорные рецепторы.
Рецепторы (от лат. receptum - принимать) приспособлены воспринимать обычно какой-то один вид раздражителей (адекватный) и максимально чувствительны к нему. Например, рецепторы сетчатки реагируют на свет, но не возбуждаются при воздействии звукового раздражителя. К другим - неадекватным раздражителям - они малочувствительны. Однако неадекватные раздражители такжемогут возбудить сенсорные рецепторы. Например, механическое воздействие на глаз вызывает ощущение света, однако энергия неадекватного раздражителя должна быть во много раз больше энергии адекватного.
Б. Классификация сенсорных рецепторов. Нервная система отличается большим разнообразием рецепторов (рис. 1.8).
Рис. 1.8. Различные типы рсцепторных клеток. Стрелки указывают на участки действия раздражителей (по Г. Шсперду, 1987, с изменениями). 1,2- соматосенсорные рецепторы; 3 - мышечный рецептор; 4 - обонятельный рецептор; 5 - слуховой и вестибулярный рецепторы; 6 - вкусовой рецептор; 7 - зрительный рецептор 1. Согласно структурно-функциональной организации, различают первичные и вторичные рецепторы. Первичные рецепторы представляют собой чувствительные окончания дендрита сенсорного нейрона, тело нейрона расположено в спинномозговом ганглии или в ганглиях черепных нервов. В первичном рецепторераздражитель действует непосредственно на окончания дендрита сенсорного нейрона.
Вторичные рецепторы имеют специальную клетку, синаптиче-ски связанную с окончанием дендрита сенсорного нейрона. Ко вторичным рецепторам относятся фоторецепторы, слуховые, вестибулярные, вкусовые рецепторы.
2. По скорости адаптации рецепторы делят на три группы: быстро адаптирующиеся (фазные), медленно адаптирующиеся (тонические) и смешанные (фазнотонические), адаптирующиеся со средней скоростью. Примером быстро адаптирующихся рецепторов являются кожные рецепторы вибрации (тельца Пачини) и прикосновения (тельца Мейснера). К медленно адаптирующимся рецепторам относятся проприорецепторы, рецепторы растяжения легких, болевые рецепторы. Со средней скоростью адаптируются фоторецепторы сетчатки, терморецепторы кожи.
3. В зависимости от вида воспринимаемого раздражителя выделяют 5 типов рецепторов, 1) Механореиетпоры возбуждаются при механической их деформации. Они расположены в коже, внутренних органах, опорно-двигательном аппарате, слуховой и вестибулярной системах. 2) Хеморецепторы воспринимают химические изменения внешней и внутренней среды организма. Такие рецепторы есть в слизистой оболочке языка (вкусовые рецепторы), носа (обонятельные рецепторы), каротидном и аортальном тельцах, продолговатом мозге, гипоталамусе. 3) Терморецепторы воспринимают изменения температуры. Они подразделяются на тепловые и холодовые рецепторы, находятся в коже, сосудах, внутренних органах, в продолговатом и спинном мозге, в среднем мозге и гипоталамусе. 4) Фоторецеппюры воспринимают световую (электромагнитную) энергию, расположены в сетчатке глаза. 5) Ноцицепторы (болевые рецепторы) отвечают на механические, термические, химические (тетании, брадикинин, К + . Н* и др.) раздражители. Болевые стимулы воспринимаются свободными нервными окончаниями.
4. По расположению в организме выделяют экстеро- и интеро-рецепторы. К экстерорецепторам относятся рецепторы кожи, видимых слизистых оболочек и органов чувств. Интерореиепторами являются рецепторы внутренних органов (висцерорецепторы), сосудов и ЦНС; проприорецепторы - рецепторы опорно-двигательного аппарата и вестибулярные рецепторы.
5. С психофизиологической точки зрения рецепторы подразделяются в соответствии с органами чувств и формируемыми ощущениями на зрительные, слуховые, тактильные, вкусовые и обонятельные.
6. В зависимости от степени специфичности рецепторов, т.е. способности отвечать на один или более видов раздражителей, выделяют мономодальные рецепторы, воспринимающие один вид раздражителей и полимодальные, воспринимающие два-три вида раздражителей. Мономодальными рецепторами являются зрительные, вкусовые, обонятельные. Различия в чувствительности к адекватным и неадекватным раздражителям у полимодальных рецепторов выражены меньше, чем у мономодальных. Пример полимодальных рецепторов - болевые рецепторы.
В. Свойства рецепторов. 1. Высокая возбудимость рецепторов. Например, волосковые рецепторы внутреннего уха способны обнаружить движение мембраны кортиева органа, равное диаметру атома водорода; для возбуждения фоторецептора сетчатки достаточно одного кванта света, обонятельного рецептора - одной молекулы пахучего вещества. Возбудимость различных рецепторов неодинакова. У висцерорецепторов она ниже, чем у экстерорецепторов. Высокая возбудимость рецепторов обеспечивает восприятие малейших изменений внутренней и внешней среды, что необходимо для надежной регуляции функций внутренних органов и приспособления организма в окружающей среде.
2. Спонтанная активность рецепторов. Многие виды рецепторов (фото-, фоно-, вестибуло-, термо-, хемо- и проприорецепторы) генерируют в окончании сенсорного нейрона потенциалы действия (ПД) в отсутствие раздражителя. Это связано со спонтанным колебанием мембранного потенциала в рецепторе, которое периодически достигает критического уровня деполяризации, что приводит к генерации ПД в нервном волокне. Возбудимость таких рецепторов выше, чем возбудимость рецепторов без фоновой активности. Значение фоновой активности рецепторов заключается в том, что она участвует в поддержании тонуса нервных центров в условиях физиологического покоя и бодрствующего состояния организма.
3. Адаптация рецепторов - уменьшение их возбудимости при длительном действии раздражителя. Процессы адаптации, формирующиеся на разных этапах преобразования информации, приводят к снижению амплитуды рецепторного потенциала и, как следствие, уменьшается частота импульсации сенсорного нейрона. Механизм адаптации рецепторов изучен недостаточно. Одним из факторов является накопление Са 2+ внутри клетки при ее возбуждении, что может ингибировать каскад ферментативных реакций в рецепторе. Другим возможным механизмом действия Са 2+ является активирование Са-зависимых калиевых каналов. Выход К + через эти каналы из клетки препятствует деполяризации ее мембраны при формировании рецепторного потенциала.
Значение адаптации рецепторов заключается в том, что предотвращается избыточное поступление информации в ЦНС и устраняются неприятные ощущения.
Г. Механизм восприятия раздражителя, действующего на рецепторы. Несмотря на большое многообразие рецепторов, в каждом из них можно выделить два или три этапа преобразования энергии раздражения в нервный импульс в зависимости от структурной организации рецептора. Первый этап - возникновение рецепторного потенциала при действии раздражителя на рецептор - одинаковый для всех рецепторов.
Действие раздражителя способно вызывать конформационные изменения рецепторного белка. На данном этапе во многих рецепторах происходит усиление сигнала, так как энергия формирующегося рецепторного потенциала оказывается многократно (например, в фоторецепторе в 10 5 раз) больше пороговой энергии раздражения.
В рецепторах (кроме фоторецепторов) энергия раздражителя приводит к открытию натриевых каналов и появлению ионных токов, среди которых основную роль играет входящий натриевый ток. Он приводит к деполяризации мембраны рецептора. Считают, что в хеморецепторах открытие каналов связано с изменением формы (конформацией) белковых молекул, выполняющих функцию ворот, а в механореценторах - с растяжением мембраны и расширением каналов. В фоторецепторах ионы натрия проникают в темноте в клетку, при действии света натриевые каналы закрываются, что уменьшает входящий натриевый ток. В связи с этим рецепторный потенциал представлен не деполяризацией, а гиперполяризацией вследствие выхода К* из клетки при уменьшении входа На + в клетку.
Чтобы информация сенсорных раздражителей достигла ЦНС, рецепторный потенциал должен быть преобразован в ПД. В первичных и вторичных рецепторах это происходит разными способами. Второй этап в первичных и вторичных рецепторах протекает по-разному. В первичном рецепторе рецеп-торная зона является частью окончаний сенсорного нейрона. Возникший рецепторный потенциал вызывает деполяризацию в участке афферентного волокна, в котором возможно возникновение ПД. В миелиновых волокнах ПД возникает в ближайших перехватах Ранвье, в безмиелиновых - в ближайших участках, имеющих достаточную концентрацию потенциалзави-симых натриевых и калиевых каналов. Если деполяризация мембраны при этом достигает критического уровня, происходит генерация ПД (рис. 1.9), т.е. процесс возникновения возбуждения происходит в два этапа.
Во вторичных рецепторах преобразование энергии раздражения в нервный импульс происходит в три этапа. На I этапе возникает рецепторный потенциал в эпителиальной рецепторной клетке, синаптически связанной с окончанием сенсорного нейрона. Рецепторный потенциал вызывает выделение в синаптическую щель медиатора. Под влиянием медиатора на постсинаптической мембране возникает генераторный потенциал (возбуждающий постсинаптический потенциал) - это II этап. Ш этап - возникновение ПД в нервном волокне вблизи постсинаптической мембраны под влиянием генераторного потенциала.
Рис. 1.9. Типичные взаимоотношения между рецепторным потенциалом и потенциалом действия при сверхпороговом уровне рецепторного потенциала (по А. Гайтону, 1986, с изменениями). КУД - критический уровень деполяризации; РП - рецепторный потенциал; МП - мембранный потенциал После поступления афферентных импульсов от рецепторов (рефлексогенных зон) в нервный центр формируется ответная рефлекторная реакция организма на раздражения. Основные положения рефлекторного принципа регуляции сформировались на протяжении весьма длительного отрезка времени. 1.1.7. Развитие концепции рефлекса. Нервизм и нервный центр А. Основные положения рефлекторного принципа деятельности ЦНС были разработаны на протяже-нии примерно двух с половиной веков. Можно выделить пять основных этапов развития этой концепции.
Этап I - заложены основы понимания рефлекторного принципа деятельности ЦНС французским естествоиспытателем и математиком Р1Декартом (XVII в.). Р.Декарт считал, что «все вещи и явления можно объяснить естественнонаучным путем». Эта исходная позиция позволила Р.Декарту сформулировать два важных положения рефлекторной теории:
1) всякая реакция на внешнее воздействие является отраженной (впоследствии ее стали называть рефлекторной);
2) ответная реакция на раздражение осуществляется при помощи нервной системы. Однако Р. Декарт был дуалистом.
По Р.Декарту, нервы - это трубочки, по которым с огромной скоростью движутся животные духи, материальные частицы неизвестной природы, по нервам они попадают в мышцу и мышца раздувается (сокращается).
Этап II - экспериментальное обоснование материалистических представлений о рефлексе (ХVII-Х1Х вв.). В частности, было установлено, что рефлекторная реакция может осуществляться на одном метамере лягушки (сегмент спинного мозга, связанный с «кусочком тела»). Выявлено, что стимулы могут быть не только внешними, но и внутренними, установлена роль задних - чувствительных и передних - двигательных корешков спинного мозга (закон Белла - Мажанди).
Весьма активно сегментарные рефлексы изучал Ч.Шеррингтон (конец XVIII- начало XIX в.).
Этап III - победа материалистических представлений о психической деятельности (И.М.Сеченов, 60-е гг. XIX в.). Наблюдая за развитием детей, И. М. Сеченов пришел к заключению, что в основе формирования психической деятельности лежит принцип рефлекса. Это положение он выразил следующей фразой: «Все акты сознательной и бессознательной жизни по способу происхождения суть рефлексы». Таким образом, И.М.Сеченов стал на путь детерминизма в вопросах психической деятельности человека. При изучении рефлексов он обосновал приспособительный характер изменчивости рефлекса, открыл торможение рефлексов (центральное торможение; 1863), суммацию возбуждения в ЦНС (1868).
Этап IV - разработаны основы учения о высшей нервной деятельности (И.П.Павлов, начало XX в.). И.П.Павлов открыл условные рефлексы и использовал их как объективный метод изучения психической деятельности (высшей нервной деятельности - по И.П.Павлову). Он сформулировал 3 принципа рефлекторной теории:
1. Принцип детерминизма (принцип причинности), согласно которому любая рефлекторная реакция причинно обусловлена.
2. Принцип структурности, суть которого заключается в том, что каждая рефлекторная реакция осуществляется с помощью определенных структур и чем больше структурных элементов участвует в осуществлении этой реакции, тем она совершеннее.
3. Принцип единства процессов анализа и синтеза в составе рефлекторной реакции: нервная система анализирует (различает) с помощью рецепторов все действующие внешние и внутренние раздражители и на основании этого анализа формирует целостную ответную реакцию (синтез).
Этап V - создано учение о функциональных системах (П.К.Анохин, середина XXв.; см. раздел 1.5).
Б. Нервизм - это концепция, признающая ведущую роль нервной системы в регуляции функций всех органов и тканей организма (физиологический нервизм). Концепция нервизма прошла весьма длинный исторический путь своего развития. Почву для этой концепции подготовил Р.Декарт (1596-1650), выдвинувший идею о рефлекторном принципе деятельности нервной системы. Ф.Гофман (1660-1742) сформулировал гипотезу о влиянии нервов на «все перемены в здоровом и больном состоянии». Согласно У.Кулену (1712-1790), все процессы в здоровом и больном организме регулирует «нервный принцип», который проявляет свое действие через головной мозг при посредстве нервов - проводников нервной деятельности. По мнению Е.О.Мухина (1817), «все человеческое тело вообще можно, отвлекаясь, рассматривать как построенное из нервов, ибо остальные части тела, видимо, существуют вследствие нервов, как управляющих их способностями».
Велики заслуги в развитии концепции нервизма И.М.Сеченова (1829-1905) и "С.П.Боткина (1832-1889). По мнению С.П.Боткина, организм - это целостная система, деятельность которой направляется и регулируется нервной системой. Он рассматривал различные заболевания как следствие нарушения нормальных нервных соотношений - клинический нервизм. Нарушение «регуляторных нервных аппаратов» может явиться причиной ряда болезней человека, что убедительно подтверждено клиническими наблюдениями.
Выдающийся вклад в развитие концепции физиологического нервизма сделал И.П.Павлов (1849-1936). Он обосновал представление о трофическом влиянии нервной системы на органы и ткани, сформулировал принципы рефлекторной теории, доказал важную роль нервной системы в регуляции секреции желез пищеварительного тракта, открыл условные рефлексы и с их помощью разработал основы учения о высшей нервной деятельности.
В. Нервный центр - это совокупность нейронов, расположенных на различных уровнях ЦНС, достаточных для приспособительной регуляции функции органа согласно потребностям организма. Например, нейроны дыхательного центра локализуются и в спинном мозге, и в продолговатом мозге, и в мосту. Однако среди нескольких групп клеток, расположенных на различных уровнях ЦНС, обычно имеется главная часть центра. Главная часть дыхательного центра находится в продолговатом мозге и включает инспираторные и экспираторные нейроны.
Нервный центр реализует свое влияние на эффекторы либо непосредственно с помощью эфферентных импульсов соматической и вегетативной нервных систем, либо с помощью активации вы-
работки соответствующих гормонов. Кроме нервной регуляции, в организме существуют гуморальный и миогенный механизмы регуляции. Гуморальный механизм регуляции функций органов и тканей организма осуществляется с помощью гормонов, медиаторов, метаболитов и тканевых гормонов.
Сенсорные рецепторы - это специфические клетки, настроенные на восприятие различных раздражителей внешней и внутренней среды организма и обладающие высокой чувствительностью к адекватному раздражителю.
Во-первых , сенсорные рецепторы являются первым звеном в рефлекторном пути и периферической частью более сложной структуры - анализаторов. Совокупность рецепторов, стимуляция которых приводит к изменению активности каких-либо нервных структур, называют рецептивным полем. Такой структурой могут быть афферентное волокно, афферентный нейрон, нервный центр (соответственно рецептивное поле афферентного волокна, нейрона, рефлекса). Рецептивное поле рефлекса часто называют рефлексогенной зоной.
Во-вторых , это эффекторные рецепторы (циторецепторы), представляющие собой белковые структуры клеточных мембран, а также цитоплазмы и ядра, способные связывать активные химические соединения (гормоны, медиаторы, лекарства и др.) и запускать ответные реакции клетки на эти соединения. Эффекторные рецепторы имеют все клетки организма, в нейронах их особенно много на мембранах синаптических межклеточных контактов.
Классификация сенсорных рецепторов: рефлекторная дуга - рецептор - стимул.
1. В зависимости от расположения в теле и характера воспринимаемых стимулов, рецепторы делятся на три типа:
экстерорецепторы - реагируют на стимулы, поступающие из внешней среды, например, уши, глаза и т.д.
интерорецепторы - воспринимают стимулы, поступающие из внутренней среды организма, например, рецепторы сонных артерий, реагирующие на изменение кровяного давления и содержание углекислого газа в крови.
проприорецепторы - отвечают на стимулы, связанные с положением и движением частей тела и сокращением мышц.
Находясь в сознании, человек постоянно чувствует положение своих конечностей и движение суставов, пассивное или активное. Кроме того, он точно определяет сопротивление каждому своему движению. Все эти способности вместе называются проприорецепцией, так как стимуляция соответствующих рецепторов (проприорецепторов) исходит из самого тела, а не из внешней среды. Применяется также термин глубокая чувствительность, так как большая часть проприорецепторов расположена не поверхностно, а в мышцах, сухожилиях и суставах.
Благодаря проприорецепторам человек обладает чувством положения, чувством движения и чувством силы.
Чувство положения информирует о том, под каким углом находится каждый сустав, и в конечном итоге - положение всех конечностей. Чувство положения почти не подвержено адаптации.
Чувство движения - это осознание направления и скорости движения суставов. Человек воспринимает как активное движение сустава при мышечном сокращении, так и пассивное, вызванное внешними причинами. Порог восприятия движения зависит от амплитуды и от скорости изменения угла сгибания суставов.
Чувство силы - это способность оценить мышечную силу, нужную для движений или для удержания сустава в определенном положении.
Проприорецепторы расположены во внекожных структурах, главные из которых - мышцы, сухожилия и суставные сумки.
2. В зависимости от природы воспринимаемых стимулов рецепторы классифицируются следующим образом:
Механорецепторы возбуждаются при их механической деформации; расположены в коже, сосудах, внутренних органах, опорно-двигательном аппарате, слуховой и вестибулярной системах.
Хеморецепторы воспринимают химические изменения внешней и внутренней среды организма. К ним относятся вкусовые и обонятельные рецепторы, а также рецепторы, реагирующие на изменение состава крови, лимфы, межклеточной и цереброспинальной жидкости. Такие рецепторы есть в слизистой оболочке языка и носа, каротидном и аортальном тельцах, гипоталамусе и продолговатом мозге.
Терморецепторы реагируют на изменения температуры. Они подразделяются на тепловые и холодовые рецепторы и находятся в коже, слизистых оболочках, сосудах, внутренних органах, гипоталамусе, среднем, продолговатом и спинном мозге.
Фоторецепторы в сетчатке глаза воспринимают световую (электромагнитную) энергию.
Ноцицепторы - возбуждение их сопровождается болевыми ощущениями (болевые рецепторы). Раздражителями этих рецепторов являются механические, термические и химические факторы. Болевые стимулы воспринимаются свободными нервными окончаниями, которые имеются в коже, мышцах, внутренних органах, дентине, сосудах.
3. С психофизиологической точки зрения рецепторы подразделяют в соответствии с органами чувств и формируемыми ощущениями на: зрительные, слуховые, вкусовые, обонятельные, тактильные.
4. В зависимости от степени специфичности рецепторов, т.е. их способности отвечать на один или более видов раздражителей, выделяют мономодальные и полимодальные рецепторы .
В принципе каждый рецептор может отвечать не только на адекватный, но и на неадекватный раздражитель, однако чувствительность к ним разная. Рецепторы, чувствительность которых к адекватному раздражителю намного превосходит таковую к неадекватным, называются мономодальными. Мономодальность особенно характерна для экстерорецепторов (зрительных, слуховых, вкусовых и др.), но есть мономодальные и интерорецепторы например хеморецепторы каротидного синуса.
Полимодальные рецепторы приспособлены к восприятию нескольких адекватных раздражителей, например механического и температурного или механического, химического и болевого. К полимодальным рецепторам относятся, в частности, ирритантные рецепторы легких, воспринимающие как механические (частицы пыли), так и химические (пахучие вещества) раздражители во вдыхаемом воздухе. Разница в чувствительности к адекватным и неадекватным раздражителям у полимодальных рецепторов выражена меньше, чем у мономодальных.
5. По скорости адаптации рецепторы делят на три группы:
1) быстро адаптирующиеся (фазные). Рецепторы вибрации и прикосновения кожи.
2) медленно адаптирующиеся (тонические). Проприорецепторы, рецепторы растяжения легких, часть болевых рецепторов.
3) смешанные (фазотонические), адаптирующиеся со средней скоростью. Фоторецепторы сетчатки, терморецепторы кожи.
Свойства рецепторов
Основные свойства рецепторов - чувствительность и способность к различению. Эти свойства обеспечиваются особыми структурными и функциональными приспособлениями:
1. Параллельные сенсорные клетки с различными порогами возбуждения - клетки с низким порогом возбуждаются под действием слабых стимулов, а по мере возрастания силы раздражителя в отходящем от клетки нервном волокне частота импульсов увеличивается. В определенной точке наступает насыщение, и дальнейшее усиление импульса уже не повышает частоту импульсов, однако при этом возбуждаются сенсорные клетки с более высоким порогом чувствительности и начинают посылать импульсы, частота которых пропорциональна силе действующего стимула. Таким образом диапазон эффективного восприятия расширяется.
2. Адаптация - при длительном воздействии сильного раздражителя большинство рецепторов вначале возбуждает в сенсорном нейроне импульсы с большой частотой, но постепенно частота их снижается. Это ослабление ответа во времени называется адаптацией. Скорость наступления и степень адаптации рецепторной клетки зависят от ее функции.
Различают медленно адаптирующиеся рецепторы и быстро адаптирующиеся рецепторы. Значение адаптации в том, что при отсутствии изменений в окружающей среде клетки находятся в покое, что предотвращает перегрузку нервной системы ненужной информацией.
3. Конвергенция и суммация. В некоторых случаях выходные пути от нескольких рецепторных клеток сходятся, т.е. конвергируют, к одному сенсорному нейрону. Воздействие стимула на одну из этих клеток не могло бы вызвать ответ в сенсорном нейроне, а одновременная стимуляция нескольких клеток дает достаточный суммарный эффект. Это явление называется суммацией.
4. Обратная связь в регуляции рецепторов. В некоторых органах чувств порог чувствительности может изменяться под действием импульсов, поступающих из центральной нервной системы. Во многих случаях эта регуляция осуществляется по принципу обратной связи с рецептором и вызывает изменения во вспомогательных структурах, благодаря чему рецепторная клетка функционирует в ином диапазоне величин стимула.
5. Латеральное торможение - оно состоит в том, что соседние сенсорные клетки, возбуждаясь, оказывают друг на друга тормозящее действие. Латеральное торможение усиливает контраст между двумя соседними участками, различающимися по интенсивности стимула.
1.2.1. Структурнофункциональная характеристика сенсорных рецепторов
Свойства сенсорных рецепторов. Возбудимость рецепторов очень высока, она превышает чувствительность новейших технических аппаратов, регистрирующих соответствующие сигналы. В частности, для возбуждения фоторецептора сетчатки глаза достаточно 1-2 квантов света, для обонятельного рецептора - одной молекулы пахучего вещества. Однако возбудимость висцерорецепторов ниже, чем экстерорецепторов. Низкую возбудимость имеют болевые рецепторы, приспособленные к ответу на действие повреждающих раздражителей.
Адаптация рецепторов - это уменьшение их возбудимости при длительном действии раздражителя, выражающееся в снижении амплитуды РП и, как следствие, частоты импульсации в афферентном нервном волокне. На начальном этапе действия раздражителей важную роль в адаптации рецепторов могут играть их вспомогательные структуры. Например, быстрая адаптация рецепторов вибрации (тельца Пачини) обусловлена тем, что их капсула пропускает к нервному окончанию только быстро изменяющиеся параметры раздражителя и «отфильтровывает» его статические составляющие. Следует заметить, что термин «темновая адаптация» фоторецепторов означает повышение их возбудимости. Одним из механизмов адаптации рецепторов является накопление Са 2+ в нем при возбуждении, что активирует Са 2+ -зависимые калиевые каналы; выход К + через эти каналы из клетки препятствует деполяризации ее мембраны и, следовательно, формированию РП. Обнаружены биохимические реакции, блокирующие формирование РП. Значение адаптации рецепторов заключается в том, что она защищает организм от избыточного потока импульсов, иногда - от неприятных ощущений.
Спонтанная активность некоторых рецепторов (фоно-, вестибуло-, термо-, хемо- и проприорецепторы) без действия на них раздражителя, что связано с проницаемостью клеточной мембраны для ионов, приводящей периодически к снижению ПП до КП и генерации ПД в нервном волокне. Возбудимость таких рецепторов выше, чем рецепторов без фоновой активности, - даже слабый раздражитель способен значительно повысить частоту импульсации нейрона. Фоновая активность рецепторов в условиях физиологического покоя участвует в поддержании тонуса ЦНС и бодрствующего состояния организма.
Функцией сенсорных рецепторов (лат. sensus -чувство, receptum -принимать) является восприятие раздражителей - изменение внешней и внутренней среды организма. Это осуществляется с помощью преобразования энергии раздражения в РП, который обеспечивает возникновение нервных импульсов.
Каждый вид рецепторов в процессе эволюции приспособлен к восприятию одного или нескольких видов раздражителей. Такие раздражители называются адекватными . К ним рецепторы имеют наибольшую чувствительность (например, рецепторы сетчатки глаза возбуждаются при действии 1-2 квантов световой энергии). К другим - неадекватным раздражителям - рецепторы малочувствительны. Неадекватные раздражители также могут возбудить сенсорные рецепторы, однако энергия этих раздражителей должна быть в миллионы и миллиарды раз больше энергии адекватных. Сенсорные рецепторы являются первым звеном в рефлекторном пути и периферической частью сенсорных систем.
Классификация сенсорных рецепторов проводится по нескольким критериям (рис. 12).
Рис. 12. Классификация рецепторов на первичные и вторичные. У вторичных рецепторов имеется рецепторная клетка, к которой подходят афферентные окончания чувствительного нейрона (Агаджанян, 2007).
По структурно - функциональной организации различают первичные и вторичные рецепторы.
Первичные рецепторы представляют собой чувствительные окончания дендрита афферентного нейрона. К ним относятся обонятельные, тактильные, температурные, болевые рецепторы и проприорецепторы. Тело нейрона расположено в спинальных ганглиях или в ганглиях черепных нервов.
Вторичные рецепторы имеют специальную клетку, синаптически связанную с окончанием дендрита сенсорного нейрона. К вторичным рецепторам относятся вкусовые, фото (зрительные)-, фоно (слуховые)- и вестибулорецепторы.
По скорости адаптации различают быстро адаптирующиеся (фазные), медленно адаптирующиеся (тонические) и смешанные (фазнотонические) рецепторы, адаптирующиеся со средней скоростью. Примером быстро адаптирующихся рецепторов являются рецепторы вибрации (тельца Пачини) и прикосновения (тельца Мейснера) кожи. К медленно адаптирующимся рецепторам относятся проприорецепторы, часть болевых рецепторов, механорецепторы легких. Со средней скоростью адаптируются фоторецепторы сетчатки глаза, терморецепторы кожи.
В зависимости от вида воспринимаемого раздражителя выделяют четыре типа рецепторов, а именно: хеморецепторы - вкусовые и обонятельные рецепторы, часть сосудистых и тканевых рецепторов (реагирующих на изменение химического состава крови, лимфы, межклеточной жидкости) - имеются в гипоталамусе (например, в пищевом центре) и продолговатом мозге (дыхательный центр); механорецепторы - расположены в коже и слизистых оболочках, опорнодвигательном аппарате, сосудах, внутренних органах, слуховой, вестибулярной и тактильной сенсорных системах; терморецепторы (их подразделяют на тепловые и холодовые) - находятся в коже, сосудах, внутренних органах, различных отделах ЦНС (гипоталамусе, среднем, продолговатом и спинном мозге); фоторецепторы - расположены в сетчатке глаза, воспринимают световую (электромагнитную) энергию.
В зависимости от способности воспринимать один или более видов раздражителей выделяют моносенсорные (обладают максимальной чувствительностью к одному виду раздражителей, например рецепторы сетчатки) и полисенсорные (воспринимают несколько адекватных раздражителей, например механический и температурный или механический, химический и болевой) рецепторы. Примером являются ирритантные рецепторы легких, болевые рецепторы.
По расположению в организме рецепторы делят на экстеро- и интерорецепторы . К интерорецепторам относятся рецепторы внутренних органов (висцерорецепторы), сосудов и ЦНС. Разновидностью интерорецепторов являются рецепторы опорно-двигательного аппарата (проприорецепторы) и вестибулярные рецепторы. К экстерорецепторам относятся рецепторы кожи, видимых слизистых оболочек (например, слизистой рта) и органов чувств: зрительные, слуховые, вкусовые, терморецепторы, обонятельные.
По ощущениям рецепторы подразделяют на зрительные, слуховые, вкусовые, обонятельные терморецепторы, тактильные, болевые (ноцицепторы) - это свободные нервные окончания, которые имеются в зубах, коже, мышцах, сосудах, внутренних органах. Они возбуждаются при действии механических, термических и химических (гистамин, брадикинин, К + , Н + и др.) раздражителей.
Механизм возбуждения рецепторов (рис. 13).

Рис. 13. Механизм возникновения и проведения сигнала от рецепторной клетки (Чеснокова, 2007)
При действии адекватного раздражителя в первичном рецепторе возникает рецепторный потенциал (РП), представляющий собой деполяризацию клеточной мембраны обычно вследствие движения ионов Nа + внутрь клетки. РП - это локальный потенциал, он является раздражителем нервного окончания (за счет своего электрического поля) и обеспечивает возникновение ПД в мякотных волокнах - в первом перехвате Ранвье, в безмякотных - в непосредственной близости от рецептора.
Во вторичных рецепторах при действии раздражителя сначала также возникаем РП в рецепторной клетке вследствие движения Nа + в клетку (вкусовые рецепторы) или К + (слуховые и вестибулярные рецепторы).
Под влиянием РП в синаптическую щель выделяется медиатор, который, действуя на постсинаптическую мембрану, обеспечивает формирование генераторного потенциала ГП (тоже локальный).
Последний является раздражителем (электрическое поле), обеспечивающим возникновение ПД в нервном окончании, как и в окончаниях с первичными рецепторами.
Зависимость частоты ПД в афферентном нервном волокне от величины РП показана на рис. 14.

Рис. 14. Типичные взаимоотношения между амплитудой РП и частотой ПД, возникающих в эфферентном нервном волокне при сверхпороговых уровнях РП (Гайтон, 2008)